Що таке рухова одиниця. Класифікація рухових одиниць. поперечнополосата серцевий м'яз
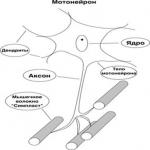
Основним морфо-функціональним елементом нервово-м'язового апарату скелетних м'язів є рухова одиниця (ДЕ). Вона включає мотонейрон спинного мозку з иннервируемой його аксоном м'язовими волокнами. Усередині м'язи цей аксон утворює кілька кінцевих гілочок. Кожна така гілочка утворює контакт - нервово-м'язовий синапс на окремому м'язовому волокні. Нервові імпульси, що йдуть від мотонейрона, викликають скорочення певної групи м'язових волокон. Рухові одиниці дрібних м'язів, які здійснюють тонкі руху (м'язи очі, кисті), містять невелику кількість м'язових волокон. У великих їх в сотні разів більше. Все ДЕ залежно від функціональних особливостей діляться на 3 групи:
I. Повільні неутомляемие. Вони утворені "червоними" м'язовими волокнами, в яких менше міофібрил. Швидкість скорочення і сила цих волокон відносно невеликі, але вони мало стомлюваність. Тому їх відносять до тонічним. Регуляція скорочень таких волокон здійснюється невеликою кількістю мотонейронів, аксони яких мають мало кінцевих гілочок. Приклад - камбаловидная м'яз.
IIВ. Швидкі, легко стомлюються. М'язові волокна містять багато міофібрил і називаються "білими". Швидко скорочуються і розвивають більшу силу, але швидко втомлюються. Тому їх називають фазними. Мотонейрони цих ДЕ найбільші, мають товстий аксон з численними кінцевими гілочками. Вони генерують нервові імпульси великої частоти. М'язи очі.
IIA. Швидкі, стійкі до стомлення. Займають проміжне положення.
Фізіологія гладких м'язів
Гладкі м'язи є в стінках більшості органів травлення, судин, вивідних проток різних залоз, сечовивідної системи. Вони є мимовільними і забезпечують перистальтику органів травлення і сечовивідної системи, підтримка тонусу судин. На відміну від скелетних, гладкі м'язи утворені клітинами частіше веретеноподібної форми і невеликих розмірів, що не мають поперечної смугастість. Останнє пов'язано з тим, що скорочувальної апарат не володіє впорядкованою будовою. Міофібрили складаються з тонких ниток актину, які йдуть в різних напрямках і прикріплюються до різних ділянок сарколеми. Міозіновие протофібрілли розташований поруч з актиновими. Елементи саркоплазматичного ретикулума не утворюють систему трубочок. Окремі м'язові клітини з'єднуються між собою контактами з низьким електричним опором - Нексус, що забезпечує поширення збудження по всій гладком'язової структурі. Збудливість і провідність гладких м'язів нижче, ніж скелетних.
Мембранний потенціал становить 40-60 мВ, так як мембрана ГМК має відносно високу проникність для іонів натрію. Причому у багатьох гладких м'язів МП не постійний. Він періодично зменшується і знову повертається до вихідного рівня. Такі коливання називають повільними хвилями (МВ). Коли вершина повільні хвилі досягає критичного рівня деполяризації, на ній починають генеруватися потенціали дії, що супроводжуються скороченнями (рис). МВ і ПД проводяться по гладким м'язам зі швидкістю всього від 5 до 50 см / сек. Такі гладкі м'язи називають спонтанно активними, тобто вони мають автоматией. Наприклад за рахунок такої активності відбувається перистальтика кишечника. Водії ритму кишкової перистальтики розташовані в початкових відділах відповідних кишок.
Генерація ПД в ГМК обумовлена \u200b\u200bвходом в них іонів кальцію. Механізми електромеханічного сполучення також відрізняються. Скорочення розвивається за рахунок кальцію, що входить в клітку під час ПД, що опосередковують зв'язок кальцію з укороченням міофібрил найважливіший клітинний білок - кальмодулін.
Крива скорочення також відрізняється. Латентний період, період укорочення, а особливо розслаблення значно триваліший, ніж у скелетних м'язів. Скорочення триває кілька секунд. Гладких м'язів, на відміну від скелетних властиво явище пластичного тонусу. Це здатність тривалий час знаходиться в стані скорочення без значних енерговитрат і втоми. Завдяки цій властивості підтримується форма внутрішніх органів і тонус судин. Крім того гладеньких м'язів самі є рецепторами розтягування. При їх натягу починають генеруватися ПД, що призводить до скорочення ГМК. Це явище називається миогенной механізмом регуляції скорочувальної активності.
Рух - необхідна умова розвитку і існування організму, його пристосування до навколишнього середовища. Саме рух є основою цілеспрямованої поведінки, що розкривається словами Н. А. Бернштейна: «Очевидна величезна біологічна значимість рухової діяльності організмів - майже єдиною форми здійснення не тільки взаємодії з навколишнім середовищем, а й активного впливу на це середовище, що змінює її з небайдужими для особини результатами ... ». Ще одним проявом значущості рухів є те, що в основі будь-якої професійної діяльності лежить робота м'язів.
Все різноманіття рухової діяльності здійснюється за допомогою опорно-рухового апарату. Його складають спеціалізовані анатомічні утворення: м'язи, скелет і центральна нервова система.
В опорно-руховому апараті з певною мірою умовності виділяють пасивну частину - скелет і активну частину - м'язи.
До скелету відносяться кістки та їх сполуки (Наприклад, суглоби).
скелет служить опорою внутрішнім органам, місцем прикріплення м'язів, захищає внутрішні органи від зовнішніх механічних пошкоджень. У кістках скелета розташований кістковий мозок - орган кровотворення. До складу кісток входить велика кількість мінеральних речовин (найбільшою мірою представлені кальцій, натрій, магній, фосфор, хлор). Кость являє собою динамічну живу тканину з високою чутливістю до різних регуляторних механізмів, до ендо- - і екзогенних впливів. Кость - не тільки орган опори, а й найважливіший учасник мінерального обміну (докладніше - в розділі Обмін речовин). Інтегральним показником метаболічної активності кісткової тканини служать тривають протягом усього життя процеси активної перебудови та оновлення кісткових структур. Ці процеси, з одного боку, є важливим механізмом підтримки мінерального гомеостазу, з іншого - забезпечують структурну адаптацію кістки до мінливих умов функціонування, що особливо значимо в зв'язку з регулярними заняттями фізичною культурою і спортом. В основі постійно протікають процесів кісткової перебудови лежить активність кісткових клітин - остеобластів і остеокластів.
м'язи за рахунок здатності скорочуватися надають руху окремі частини тіла, а також забезпечують підтримку заданої пози. М'язове скорочення супроводжується виробленням великої кількості тепла, а значить, працюють м'язи беруть участь в теплоутворення. Добре розвинені м'язи є чудовим захистом внутрішніх органів, судин і нервів.
Кістки і м'язи, як по масі, так і за обсягом становлять значну частину всього організму, в їх співвідношенні є істотні статеві відмінності. М'язова маса дорослого чоловіка - від 35 до 50% (в залежності від того, наскільки розвинені м'язи) від загальної маси тіла, жінки - приблизно 32-36%. У спортсменів, що спеціалізуються в силових видах спорту, м'язова маса може досягати 50-55%, а у культуристів - 60-70% загальної маси тіла. На частку кісток припадає 18% від маси тіла у чоловіків і 16% у жінок.
У людини розрізняють три види м'язів:
поперечносмугасті скелетні м'язи;
поперечнополосата серцевий м'яз;
гладкі м'язи внутрішніх органів, шкіри, судин.
гладкі м'язиподіляються на тонічні(Не здатні розвивати «швидкі» скорочення, в сфінктерах порожнистих органів) і фазно-тонічні (Які діляться на що володіють автоматией, Тобто здатністю до спонтанної генерації фазних скорочень. Прикладом можуть бути м'язи органів шлунково-кишкового тракту і сечоводів, і що не володіють цією властивістю - м'язовий шар артерій, насінних протоків, м'яз райдужки ока, вони скорочуються під впливом імпульсів вегетативної нервової системи. Рухова іннервація гладких м'язів здійснюється відростками клітин вегетативної нервової системи, чутлива - відростками клітин спінальних гангліїв. Як правило, скорочення гладкої мускулатури не може бути викликано довільно, в регуляції її скорочень не бере кора мозку. Функція гладких м'язів полягає в тому, щоб підтримувати тривале напруження, при цьому вони витрачають в 5 - 10 разів менше АТФ, ніж знадобилося б для виконання такої ж завдання скелетної м'язі.
Гладкі м'язи забезпечують функцію порожнистих органів, Стінки яких вони утворюють. Завдяки гладким м'язам здійснюється вигнання вмісту з сечового міхура, кишок, шлунка, жовчного міхура, матки. Гладкі м'язи забезпечують сфінктерную функцію - створюють умови для зберігання певного вмісту в підлогою органі (сечі в сечовому міхурі, плода в матці). Змінюючи просвіт кровоносних судин, гладкі м'язи адаптують регіонарний кровотік до місцевих потреб в кисні і поживних речовин, беруть участь в регуляції дихання за рахунок зміни просвіту бронхіального дерева.
скелетні м'язи є активною частиною опорно-рухового апарату, забезпечуючи цілеспрямовану діяльність, в першу чергу за рахунок довільних рухів (докладніше особливості їх будови і принципів роботи розглянуті нижче).
Види м'язових волокон
М'язи складаються з м'язових волокон, що володіють різною силою, швидкістю і тривалістю скорочення, а також стомлюваністю. Ферменти в них мають різною активністю і представлені в різних ізомерних формах. Помітно відмінність в них змісту дихальних ферментів - гликолитических і окислювальних. За співвідношенням міофібрил, мітохондрій і міоглобіну розрізняють так звані білі, червоні і проміжні волокна . За функціональним особливостям м'язові волокна ділять на швидкі, повільні і проміжні . Якщо по активності АТФази м'язові волокна розрізняються досить різко, то ступінь активності дихальних ферментів варіює дуже значно, тому поряд з білими і червоними існують і проміжні волокна.
Найбільш явно м'язові волокна розрізняються особливостями молекулярної організації міозину. Серед різних його ізоформ існують дві основні - «швидка» і «повільна». При постановці гістохімічних реакцій їх розрізняють по АТФазной активності. З цими властивостями корелює і активність дихальних ферментів. зазвичай в швидких волокнах (FF-волокна - швидко скорочуються,fast twitch fibres), переважають гліколітичні процеси, вони багатші глікогеном, в них менше міоглобіну, тому їх називають також білими. В повільних волокнах, Що позначаються як S (ST) волокна (slow twitch fibres), навпаки, вище активність окисних ферментів, вони багатшими миоглобином, виглядають більш червоними. Вони включаються при навантаженнях в межах 20-25% від максимальної сили і відрізняються гарною витривалістю.
FT - волокна, що володіють у порівнянні з червоними волокнами невеликим вмістом міоглобіну, характеризуються високою скорочувальної швидкістю і можливістю розвивати велику силу. У порівнянні з повільними волокнами вони можуть удвічі швидше скорочуватися і розвинути в 10 разів більшу силу. FT-волокна, в свою чергу, поділяються на FTO-і FTG-волокна. Істотні відмінності між перерахованими типами м'язових волокон визначається способом отримання енергії (рис.2.1).
Мал. 2.1Разлічія енергозабезпечення у м'язових волокон різних типів (По http://medi.ru/doc/g740203.htm).
Отримання енергії в FTO-волокнах відбувається так само, як і в ST-волокнах, переважно шляхом окисного фосфорилювання. У зв'язку з тим, що цей процес розкладання протікає щодо економічно (на кожну молекулу глюкози при розкладанні м'язовогоглікогену для отримання енергії накопичується 39 енергетичних фосфатних сполук), FTO-волокна мають також відносно високу опірність стомлюваності. Накопичення енергії в FTG-волокнах відбувається переважно шляхом гліколізу, т. Е. Глюкоза в відсутності кисню розпадається до ще відносно багатого енергією лактату. У зв'язку з тим, що цей процес розпаду неекономічний (на кожну молекулу глюкози для отримання енергії накопичується лише 3 енергетичних фосфатних з'єднання), FTG-волокна відносно швидко втомлюються, але, тим не менше вони здатні розвинути велику силу і, як правило, включаються при субмаксимальних і максимальних м'язових скороченнях.
рухові одиниці
Основним морфофункциональним елементом нервово-м'язового апарату скелетних м'язів є рухова одиниця - ДЕ(Рис.2.2.).

Рис 2.2. рухова одиниця
ДЕ включає мотонейрони спинного мозку з иннервируемой його аксоном м'язовими волокнами. Усередині м'язи цей аксон утворює кілька кінцевих гілочок. Кожна така гілочка утворює контакт - нервово-м'язовий синапс на окремому м'язовому волокні. Нервові імпульси, що йдуть від мотонейрона, викликають скорочення певної групи м'язових волокон. ДЕ дрібних м'язів, які здійснюють тонкі руху (м'язи очі, кисті), містять невелику кількість м'язових волокон. У великих м'язах їх в сотні разів більше.
ДЕ активізуються за законом "все або нічого". Таким чином, якщо від тіла мотонейрона переднього рогу спинного мозку надсилається по нервових шляхах імпульс, то на нього реагують або все м'язові волокна ДЕ, або жодного. Для біцепса це означає наступне: при нервовому імпульсі необхідної сили коротшають всі скорочувальні елементи (міофібрили) всіх (приблизно 1500) м'язових волокон відповідної ДЕ.
Все ДЕ залежно від функціональних особливостей діляться на 3 групи:
I. повільні неутомляемие. Вони утворені "червоними" м'язовими волокнами, в яких менше міофібрил. Швидкість скорочення і сила цих волокон відносно невеликі, але вони мало стомлюваність, тому ці волокна відносять до тонічним. Регуляція скорочень таких, волокон здійснюється невеликою кількістю мотонейронів, аксони яких мають мало кінцевих гілочок. Приклад - камбаловидная м'яз.
II В. Швидкі, легко стомлюються. М'язові волокна містять багато міофібрил і називаються "білими". Швидко скорочуються і розвивають більшу силу, але швидко втомлюються. Тому їх називають фазними. Мотонейрони цих ДЕ найбільші, мають товстий аксон з численними кінцевими гілочками. Вони генерують нервові імпульси великої частоти. Напр., М'язи ока.
II А. Швидкі, стійкі до стомлення (Проміжні).
Всі м'язові волокна однієї ДЕ відносяться до одного і того ж типу волокон (FT- або ST-волокна).
М'язи, задіяні у виконанні дуже точних і диференційованих рухів (наприклад, м'язи очей або пальців руки), складаються зазвичай з великої кількості ДЕ (від 1500 до 3000). Такі ДЕ мають невелику кількість м'язових волокон (від 8 до 50). М'язи, що виконують відносно менш точні рухи (наприклад, великі м'язи кінцівок), мають істотно меншою кількістю ДЕ, але в їх склад включено велику кількість волокон (від 600 до 2000).
В середньому людина має приблизно 40% повільних і 60% швидких волокон. Але це середня величина (по всій скелетної мускулатури), м'язи ж виконують різні функції. Кількісний та якісний склад м'язів неоднорідний, в них входить різне число рухових одиниць, співвідношення типів яких також по-різному ( композиція м'язів). У зв'язку з цим скоротливі здатності різних м'язів неоднакові. Зовнішні м'язи ока, які обертають очне яблуко, розвивають максимальну напругу за одне скорочення тривалістю всього 7.5 мс, камбалообразних - антигравітаційна м'яз нижньої кінцівки, дуже повільно розвиває максимальну напругу протягом 100 мс. М'язи, що виконують велику статичну роботу (камбаловидная м'яз), часто мають більшу кількість повільних ST-волокон, а м'язи, які вчиняють переважно динамічні руху (біцепс), мають велику кількість FT-волокон.
Основні властивості м'язових волокон (отже, і рухових одиниць - ДЕ, до складу яких вони входять), які визначаються також властивостями мотонейронів, представлені в таблиці 1.
Величина скорочення (сила м'язи) залежить від морфологічних властивостей і фізіологічного стану м'язи:
1. Вихідною довжини м'язи (довгі спокою). Сила м'язового скорочення залежить від вихідної довжини м'язи або довжини спокою. Чим сильніше м'яз розтягнута в спокої, тим сильніше скорочення (закон Франка-Старлінг).
2. Діаметра м'язи або поперечного перерізу. Виділяють два діаметра:
а) анатомічний діаметр - поперечний переріз м'язів.
б) фізіологічний діаметр - перпендикулярний переріз кожного м'язового волокна. Чим більше фізіологічне перетин, тим більшою силою володіє м'яз.
Сила м'яза вимірюється вагою максимального вантажу піднятого на висоту або максимальною напругою, яке вона здатна розвинути в умовах ізометричного скорочення. Вимірюється в кілограмах або ньютонах. Методика вимірювання сили м'язи називається динамометрия.
Виділяють два види сили м'язи:
1. Абсолютна сила - відношення максимальної сили до фізіологічного діаметру.
2. Відносна сила - відношення максимальної сили до анатомічного діаметру.
При скороченні м'яз здатний виконувати роботу. Робота м'язи вимірюється твором піднятого вантажу на величину укорочення.
Робота м'язи характеризується потужністю. Потужність м'язи визначається величиною роботи в одиницю часу і вимірюється у ВАТ.
Найбільша робота і потужність досягається при середніх навантаженнях.
Мотонейрон з групою іннервіруємих їм м'язових волокон становить рухову одиницю. Аксон мотонейронів може гілки і іннервувати групу м'язових волокон. Так, один аксон може іннервувати від 10 до 3000 м'язових волокон.
Розрізняють рухові одиниці за будовою і функціями.
За будовою рухові одиниці поділяються на:
1. Малі рухові одиниці, які мають малий мотонейрон і тонкий аксон, здатний іннервувати 10-12 м'язових волокон. Наприклад, м'язи обличчя, м'язи пальців рук.
2. Великі рухові одиниці представлені великим тілом мотонейрона, товстим аксонів, який здатний іннервувати більше 1000 м'язових волокон. Наприклад, чотириглавий м'яз.
За функціональним значенням рухові одиниці поділяються на:
1. Повільні рухові одиниці. Вони включають малі рухові одиниці, є легко збудливими, характеризуються невисокою швидкістю поширення збудження, в роботу включаються першими, але при цьому вони практично не стомлюваність.
2. Швидкі рухові одиниці. Вони складаються з великих рухових одиниць, погано збудливі, володіють великою швидкістю проведення збудження. Мають високу силою і швидкістю відповіді. Наприклад, м'язи боксера.
Ці особливості рухових одиниць зумовлені низкою властивостей.
М'язові волокна, які входять в рухові одиниці, мають подібні властивості і відмінності. Так, повільні м'язові волокна володіють:
1. Багатою капілярної мережею.
4. У них міститься багато жирів.
Завдяки цим особливостям ці м'язові волокна володіють високою витривалістю, здатні до невеликих за силою скорочень, але тривалим за часом.
Відмінні риси швидких м'язових волокон:
2. Мають більшою швидкістю і силою скорочення.
У зв'язку з цими особливостями швидкі м'язові волокна швидка стомлюваність, але володіють великою силою і високою швидкістю відповіді.
За морфофункциональним властивостями рухові одиниці поділяються на 3 типи:
1. Повільні неутомляемие ДЕ. Мотонейрони мають найнижчий поріг активації, здатні підтримувати стійку частоту розрядів протягом десятків хвилин (тобто неутомляемость). Аксони мають невелику товщину, низькою швидкістю проведення збудження, іннервують невелику групу м'язових волокон. М'язові волокна розвивають невелику силу при скороченні в зв'язку з наявністю в них найменшої кількості скорочувальних білків - міофібрил. Це так звані «червоні волокна» (колір обумовлений хорошим розвитком капілярної мережі і невеликою кількістю міофібрил). Швидкість скорочення цих волокон в 1,5 - 2 рази менше, ніж швидких. Вони неутомляемость завдяки добре розвиненою капілярної мережі, великій кількості мітохондрій і високої активності окислювальних ферментів.
2. Швидкі, легко стомлюються ДЕ. Мають найбільш великий мотонейрон, що володіє найбільш високим порогом збудження, не здатні протягом тривалого часу підтримувати стійку частоту розрядів (стомлюються). Аксони товсті, з великою швидкістю проведення нервових імпульсів, іннервує багато м'язових волокон. М'язові волокна містять велику кількість міофібрил, тому при скороченні розвивають більшу силу. Завдяки високій активності ферментів швидкість скорочення висока. Ці волокна швидко втомлюються, тому що містять менше, в порівнянні з повільними, мітохондрій і оточені меншою кількістю капілярів.
3. Швидкі, стійкі до стомлення. Сильні, швидко скорочуються волокна, що володіють великою витривалістю завдяки можливості використання аеробних і анаеробних процесів отримання енергії. Волокна 2 і 3 типів називаються «білими волокнами» через великий вміст міофібрил і низького - міоглобіну.
Порівняння повільних і швидких м'язових волокон
Скелетний м'яз людини складається з волокон 3 типів, однак їх співвідношення може значно відрізнятися в залежності від функції м'язи, а також вродженої та набутої індивідуальності. Чим більше в м'язах білих волокон, тим краще людина пристосований до виконання роботи, що вимагає великої швидкості і сили. Переважання червоних волокон забезпечує витривалість при виконанні тривалої роботи.
Будова скелетного м'яза
Скелетний м'яз складається з безлічі м'язових волокон, які розташовані пучками в загальному сполучнотканинному футлярі і кріпляться до сухожиль, пов'язаним зі скелетом. Кожне м'язове волокно - це тонке (від 10 до 100 мкм) витягнуте в довжину (від 5 до 400мм) багатоядерне освіту - симпласт.
Мембрани м'язового волокна подібна за будовою з нервовою, але вона дає регулярні Т-образні впячивания. Всередині м'язового волокна паралельно мембрані розташовується розгалужена замкнута система трубочок - саркоплазматический ретикулум - внутрішньоклітинний депо Ca 2+. Т-система і прилеглий до неї СР - апарат передачі збудження з мембрани м'язового волокна на скоротливі структури (міофібрили). У саркоплазме м'язового волокна можна побачити поперечні чергуються світлі і темні ділянки - відповідно, J- (ізотропні) і А- (анізотропні) диски. У сусідніх миофибриллах однойменні диски розташовані на одному рівні, що надає волокну смугастість.Комплекс з одного темного і двох прилеглих до нього половин світлих дисків, обмежених поперечними Z-пластинками, називають саркомером.
Кожна миофибрилла складається їх безлічі паралельно лежачих товстих (миозинових) і тонких (Актинові) білкових ниток - миофиламентов. За перетину волокна товсті і тонкі нитки розташовуються високоорганізованих в вузлах гексагональної решітки. Кожна товста нитка оточена шістьма тонкими, кожна з тонких ниток частково входить в оточення трьох сусідніх товстих. Міозіновие нитки мають відходять від них поперечні виступи з головками, що складаються приблизно з 150 молекул міозину. Актинового нитка складається з двох закручених одна навколо іншої ланцюжків (подібно скрученим ниткам бус) молекул актину. На нитках актину розташовані молекули тропонина, а в жолобках між двома нитками актину лежать нитки тропомиозина.
Механізм скорочення м'язового волокна
У 1954 р Г.Хакслі і Н.Хенсон виявили, що Актинові і міозіновие філаменти не змінюють своєї довжини при вкороченні або подовженні саркомера і вивели теорію ковзання ниток: М'язове скорочення відбувається при послідовному зв'язуванні декількох центрів міозіновой головки поперечного містка з певними ділянками на Актинові філаментах.
У покояться м'язових волокнах молекули тропомиозина в спокої розташовуються так, що запобігають прикріплення поперечних містків міозину до Актинові ниток (м'яз розслаблений).
Виникає в області аксо-соматичного синапсу ПД поширюється по системі Т-трубочок вглиб волокна, викликаючи деполяризацію цистерн саркоплазматичного ретикулума (депо Са 2+). При активації мембрани СР відбувається відкриття Са-каналів і вихід Са 2+ по концентраційному градієнту.
При підвищенні в міоплазми концентрації іонів Са 2+ він з'єднується з тропонином, останній конформируется і відсуває нитка тропомиозина, відкриваючи для міозіновой головки можливість з'єднання з актином. З'єднання головки приводить до різкого «згинання» містка і переміщенню нитки актину на 1 крок (20 нм або 1% довжини актину) до середини саркомера з подальшим розривом містка.
При відсутності повторного порушення концентрація Са 2+ завдяки роботі Са-насоса падає. Тому Са 2+ від'єднується від тропонина і тропомиозин знову блокує актин. При цьому на одне робоче рух одного містка витрачається енергія 1 молекули АТФ, ще однією - на повернення 2 іонів Са 2+ в цистерни.
Все це призводить до розслаблення м'язи аж до моменту приходу чергового потоку нервових імпульсів, коли описаний вище процес повторюється.
Сукупність процесів, які обумовлюють розподіл ПД вглиб м'язового волокна, вихід іонів Са 2+ з саркоплазматичного ретикулума, взаємодія скорочувальних білків і вкорочення м'язового волокна називається електромеханічним сполученням.
Механіка м'язи. Фізичні властивості і режими м'язових скорочень
Фізичні властивості скелетних м'язів
1. розтяжність- здатність м'яза змінювати свою довжину під дією розтягує її сили.
2. еластичність - здатність м'яза приймати свою первісну довжину після припинення дії розтягує або деформуючий сили.
3. сила м'язи. Вона визначається максимальним вантажем, який м'яз в змозі підняти. Питома сила - максимальний вантаж, який м'яз в змозі підняти, ділять на кількість квадратних сантиметрів її фізіологічного поперечного перерізу.
4. Здатність м'язи здійснювати роботу. Робота м'язи визначається твором величини піднятого вантажу на висоту підйому. Робота м'язи поступово збільшується зі збільшенням вантажу, але до певної межі, після якого збільшення вантажу призводить до зменшення роботи, т. К. Знижується висота підйому вантажу. Отже, максимальна робота м'язом проводиться при середніх величинах навантажень ( закон середніх навантажень).
Режими м'язових скорочень
Розрізняють ізотонічний, ізометричний і змішаний режими скорочення м'язів.
при фізіологічному скороченні м'яза відбувається зміна її довжини, а напруга залишається постійним. Таке скорочення відбувається в тому випадку, коли м'яз не переміщує вантаж. У природних умовах близькими до ізотонічного типу скорочень є скорочення м'язів мови.
при изометрическомскорочення довжина м'язових волокон залишається постійною, змінюється напруга м'язи. Таке скорочення м'язи можна отримати при спробі підняти непосильний тягар.
В цілому організмі скорочення м'язів ніколи не бувають чисто фізіологічним або изометрическим, вони завжди мають змішаний характер, т. Е. Відбувається зміна і довжини, і напруги м'язи. Такий режим скорочення називається ауксотоніческім якщо переважає напруга м'язи, або ауксометріческім якщо переважає вкорочення.
|
Швидкі |
повільні |
|
нейрон |
|
|
великі мотонейрони |
дрібні мотонейрони |
|
збудливість менше |
збудливість більше |
|
Діаметр аксона більше |
Діаметр аксона менше |
|
Швидкість проведення збудження більше |
Швидкість проведення збудження менше |
|
частота більше |
частота менше |
|
м'язові волокна |
|
|
Активність актомиозинового АТФази вище |
Активність актомиозинового АТФази менше |
|
Щільність упаковки актомиозинового филаментов вище |
Щільність упаковки актомиозинового филаментов менше |
|
Більш виражений саркоплазматический ретикулум (депо кальцію) |
Менш виражений саркоплазматический ретикулум (депо кальцію) |
|
Латентний період після надходження ПД менше |
Латентний період після надходження ПД більше |
|
Щільність кальцієвої помпи вище |
Щільність кальцієвої помпи менше |
|
Швидше скорочується і розслабляється |
Повільніше скорочується і розслабляється |
|
Вище активність ферментів гліколізу |
Вище активність ферментів окислення |
|
Швидше відновлення АТФ |
Відновлення АТФ повільніше, але економічніше |
|
1 моль глюкози -2-3 молей АТФ |
1 моль глюкози 36-58 молей АТФ |
|
Утворюються недоокислені субстрати, «закислення» - швидке стомлення |
стомлення менш виражено |
|
Велика щільність капілярів - більше оксигенація, більше міоглобіну |
|
|
рухова одиниця |
|
|
Менш збудлива, велика сила і швидкість скорочення, велика втомлюваність, низька витривалість |
Більш збудлива, менша сила, швидкість скорочення, мала стомлюваність, висока витривалість |
|
спринтери | |
|
У зовнішній м'язі стегна повільні волокна від 13 до 96% |
Триголовий м'яз плеча 33%, двоголова 49%, передня великогомілкова 46%, камбаловидная 84% |
Нейрофізіологічні основи методу електроміографії.
Електроміографія - це метод дослідження нервово-м'язової системи за допомогою реєстрації електричних потенціалів м'язів. Хоча вперше електроміограма (ЕМГ) була зареєстрована за допомогою телефонного пристрою Н. Е. Введенським ще в 1884 р, а в 1907 р вдалося здійснити графічну запис ЕМГ людини, інтенсивний розвиток електроміографії в якості клінічної діагностичної методики почалося в 30-40-е роки XX століття Певна затримка прогресу в цій області в порівнянні, наприклад, з розвитком електроенцефалографії, пояснюється високими вимогами до якості реєстрації і точності відтворення істинних параметрів електричних потенціалів в електроміографії. Створення високоякісних підсилювачів, що дають лінійні характеристики в діапазоні високих частот, і розробка методів катодного реєстрації, що забезпечує неспотворене відтворення високочастотних складових електричного потенціалу до діапазону 20000 Гц, привели до істотного прогресу в області клінічного застосування електроміографії
При внутрішньоклітинної реєстрації потенціал дії виглядає як позитивний пік, що складається з швидкої деполяризації, що триває близько 1 мс, швидкої реполяризації, що представляє собою повернення потенціалу майже до рівня спокою, що триває близько 2 мс; потім слідують повільна реполяризация, невелика следовая гиперполяризация і повернення потенціалу до рівня спокою. У клінічній електроміографії при позаклітинній реєстрації макроелектродом потенціал дії м'язового волокна представлений негативним піком тривалістю 1-3 мс.
Техніка відведення та реєстрації ЕМГ
Принципи техніки відведення та реєстрації ЕМГ не відрізняються від техніки електроенцефалографії, електрокардіографії і інших єлектрографических методів. Система складається з електродів, що відводять потенціали м'язи, підсилювача цих потенціалів і реєструючого пристрою. У електроміографії використовується два види електродів - поверхневі і голчасті. Поверхневі електроди являють собою металеві пластини або диски площею близько 0,2 - 1 см 2, зазвичай вмонтовані попарно в фіксують колодки, що забезпечують сталість відстаней між отводящими електродами, що важливо для оцінки амплітуди реєстрованого активності. Такі електроди накладають на шкіру над областю рухової точки м'язи. Шкіру перед накладенням електрода протирають спиртом і змочують фізіологічним розчином хлориду натрію. Електрод фіксують над м'язом за допомогою гумових смуг, манжет або лейкопластиру. При необхідності тривалого дослідження на область шкірно-електродного контакту наносять спеціальну електродний пасту, використовувану в електроенцефалографії. Великий розмір і віддаленість від м'язової тканини поверхневого електрода дозволяють реєструвати з його допомогою тільки сумарну активність м'язів, що представляє собою інтерференцію потенціалів дії багатьох сотень і навіть тисяч м'язових волокон. При великому посиленні і сильних м'язових скороченнях поверхневий електрод реєструє також активність сусідніх м'язів. Все це не дозволяє досліджувати за допомогою поверхневих електродів параметри окремих м'язових потенціалів. У одержуваної реєстрації тільки орієнтовно оцінюють частоту, періодичність і амплітуду ЕМГ. Перевага поверхневих електродів є атравматичность, відсутність ризику інфекції, простота поводження з електродами. Безболісність дослідження не накладає обмежень на кількість досліджуваних за один раз м'язів, робить цей метод кращим при обстеженні дітей, а також при фізіологічному контролі в спортивній медицині або при дослідженні із застосуванням масивних і сильних рухів.
Голчасті електроди бувають концентричними, біполярними і монополярний. У першому варіанті електрод представлений порожнистої голкою діаметром близько 0,5 мм всередині якої проходить відокремлений від неї шаром ізоляції дротяний стрижень з платини або нержавіючої сталі. Різниця потенціалів вимірюють між корпусом голки і кінчиком центрального стрижня. Іноді для збільшення локальності відведення голку ізолюють також зовні, причому неізольованою залишають тільки її еліптичну поверхню по площині зрізу. Площа відводить поверхні осьового стрижня стандартного концентричного електрода становить 0,07 мм 2 Наведені в сучасних публікаціях параметри потенціалів ЕМГ відносяться до електродів цього типу і розміру. При істотному збільшенні площі контакту відвідного електрода параметри потенціалів можуть істотно змінюватися. Це саме можна сказати до змін конструкції електрода (біполярний, монополярний, мультіелектрод). Біполярний електрод містить всередині голки два однакових ізольованих один від одного стержня, між оголеними кінчиками, яких, віддаленими одна від одної на десяті частки міліметра, вимірюють різницю потенціалів. Нарешті, для монополярних відведень використовують електроди, що представляють собою голку, ізольовану на всьому протязі, крім загостреного кінця, оголеного протягом 1-2 мм. Голчасті електроди використовують для дослідження параметрів ПД окремих ДЕ і м'язових волокон. Відведення голчастим електродом є основним в клінічній міографії, орієнтованої на діагностику первинно-м'язових і нервово-м'язових захворювань. Запис окремих ПД в ДЕ і м'язових волокон дозволяє точно оцінити тривалість, амплітуду, форму і фазность потенціалу
види відведень
Незалежно від типу електродів розрізняють два способи відведення електричної активності - моно- і біполярний. У електроміографії монополярним називається таке відведення, коли один електрод розташовується безпосередньо поблизу досліджуваної ділянки м'язів, а другий - у віддаленій від нього області (шкіра над кісткою, мочка вуха і ін.). Перевагою монополярного відведення є можливість визначити форму потенціалу досліджуваної структури і справжню фазу відхилення потенціалу. Недолік полягає в тому, що при великій відстані між електродами в запис втручаються потенціали від інших відділів м'язи або навіть від інших м'язів. Біполярний відведення - це таке відведення, при якому обидва електроди знаходяться на досить близькій і однаковій відстані від досліджуваної області м'язи. Таким є відведення за допомогою біполярних або концентричних голчастих електродів і за допомогою пари поверхневих електродів, зафіксованих в одній колодці. Біполярний відведення в малому ступені реєструє активність від віддалених джерел потенціалу, особливо при використанні голчастих електродів. Вплив на різницю потенціалів активності, що надходить від джерела на обидва електроди, призводить до спотворення форми потенціалу та неможливості визначити справжню фазу потенціалу. Проте високий ступінь локальності робить цей спосіб кращим в клінічній практиці. Оскільки відведення поверхневими електродами в будь-якому випадку реєструє интерференционную активність багатьох взаімоналагающіхся ПД ДЕ, використання такого монополярного відведення не має сенсу.
Крім електродів, різниця потенціалів яких подається на вхід підсилювача ЕМГ, на шкіру досліджуваного встановлюють поверхневий електрод заземлення, який приєднують до відповідної клеми на електродної панелі електроміографа. Різниця потенціалів від електродів подається на вхід підсилювача напруги. Підсилювач забезпечений ступінчастим перемикачем коефіцієнта посилення, що дозволяє регулювати рівень посилення в залежності від амплітуди реєстрованого активності. Посилену електричну активність виводять не тільки на осцилоскоп, але і на гучномовець, що дозволяє оцінювати електричні потенціали на слух
Загальні принципи аналізу ЕМГ і Електроміографічне семіотика.
Аналіз Електроміографічні кривої включає на першому етапі диференціацію власне електричних потенціалів м'язів від можливих артефактів і потім, на основному етапі, оцінку власне ЕМГ. Попередня оперативна оцінка здійснюється по екрану осцилографа і акустичним феноменам, які виникають при виведенні посиленою ЕМГ на гучномовець; остаточний аналіз з кількісною характеристикою ЕМГ і клінічним висновком виробляють по запису на папері або кіноплівці.
Артефактних потенціалами в ЕМГ називаються потенціали, не пов'язані власне з активністю м'язових елементів. При поверхневому відведенні артефакти можуть обумовлюватися рухом електрода внаслідок його нещільної фіксації на шкірі, що призводить до появи високоамплітудних стрибків потенціалу неправильної форми. При игольчатом відведенні аналогічні зміни потенціалу можуть виникати при дотику до електроду, сполучних проводів, при масивних рухах досліджуваного м'яза. Найбільш часто зустрічається видом перешкоди є наводка 50 Гц від пристроїв експлуатації промислового струму. Вона легко розпізнається за характерною синусоїдальної формі і постійної частоті і амплітуді. Виникнення її може бути пов'язано з великим електродним опором, що вимагає відповідної обробки голчастого електрода. При поверхневих електродах усунення наведення може бути досягнуто більш ретельної очищенням шкіри спиртом, використанням електродної пасти.
Аналіз ЕМГ включає оцінку форми, амплітуди і тривалості потенціалів дії окремих м'язових волокон і ДЕ і характеристику інтерференційної активності, що виникає при довільному м'язовому скороченні. Форма окремого коливання м'язового потенціалу може бути моно-, ди-. три- або поліфазний. Як і в електроенцефалографії, монофазним називається таке коливання, при якому крива робить відхилення в одну сторону від ізоелектричної лінії і повертається до вихідного рівня. Діфазним називається коливання, при якому крива по скоєнні відхилення в одну сторону від ізоелектричної лінії перетинає її і робить коливання в протилежній фазі; трифазне коливання здійснює відповідно три відхилення в протилежні сторони від ізоелектричної лінії. Поліфазним називається коливання, що містить чотири і більше фаз.
Стимуляційні методи в електроміографії
Крім дослідження електричної активності м'язів у спокої, при рефлекторних і довільних скороченнях, сучасна комплексна методика клінічної електроміографії включає дослідження електричних реакцій нервів і м'язів на електричну стимуляцію. Апаратура і способи реєстрації викликаної стимуляцією електричної активності ті ж, що і в звичайній електроміографії. Для стимуляції нервів і м'язів використовують електростимулятори. Стимуляцію м'язів виробляють нашкірними електродами в рухових точках, стимуляцію нервів згідно зонам їх проекції на шкіру. Стимулюючі електроди виготовляють у вигляді металевих дисків діаметром 6-8 мм, вмонтованих в металеву обойму і змочуваних фізіологічним розчином хлориду натрію. Стимуляційні методи в діагностиці нервово-м'язових захворювань вирішують такі основні завдання: 1) дослідження прямої збудливості м'язів; 2) дослідження нервово-м'язової передачі; 3) дослідження стану мотонейронів і їх аксонів; 4) дослідження стану чутливих волокон периферичних нервів. За допомогою електроміографії можна виявити, чи пов'язано зміна електричної активності з ураженням мотонейрона або синаптичних і надсег-плементарним структур.
Електромиографические дані широко використовуються для уточнення топічного діагнозу і об'єктивізації патологічних або відновних процесів. Висока чутливість цього методу, що дозволяє виявляти субклинические ураження нервової системи, робить його особливо цінним. Електроміографія широко застосовується не тільки в неврологічній практиці, але і при вивченні ураження інших систем, коли виникають вдруге обумовлені порушення рухової функції (серцево-судинні, обмінні, ендокринні захворювання).
При довільному розслабленні м'язів уловлюються тільки дуже слабкі (до 10-15 мкВ) і часті коливання биопотенциала. Рефлекторні зміни м'язового тонусу характеризуються незначним збільшенням амплітуд частих, швидких і мінливих за ритмом коливань біопотенціалів (до 50 мкВ). При довільних скороченнях м'язів реєструються інтерференційні електроміограми (з частими високовольтними біопотенціалів до 2000 мкВ).
Поразка клітин переднього рогу спинного мозку викликає зміна ЕМГ в залежності від тяжкості ушкодження, характеру перебігу захворювання і стадії його. При парезі спостерігаються урежении, ритмічні коливання зі збільшенням тривалості до 15-20 мс. Поразка переднього корінця або периферичного нерва викликає зниження амплітуди і частоти біопотенціалів, зміна форми ЕМГ-кривої. Млявий параліч проявляється "біоелектричним мовчанням".
ЕМГ однієї з м'язів руки людини в нормі. . Електроміограма при ураженні передніх рогів спинного мозку.

Питання для самостійної позааудиторної роботи студентів:
Склад рухової одиниці. Поняття моторного пулу.
Класифікація рухових одиниць.
Порівняльна характеристика швидких і повільних рухових одиниць.
Регуляція сили скорочення цілісної м'язи. Принципи «залучення» рухових одиниць, фракціонування моторного пулу, загального кінцевого шляху.
Метод електроміографії, принцип методу, медичне значення методу ЕМГ.
У зошиті практичних робіт підготувати коротку характеристику методу ЕМГ (принцип методу, необхідна апаратура, види електродів і особливості їх застосування, медичне значення методу).