Spermarakk koosneb järgmistest osakondadest. Sperma struktuur. Munaraku struktuur. Kuidas toimub viljastumine
Kõik teavad, et sperma on meeste sugurakk. Eksperdid nimetavad seda sugurakuks. Isaste sugurakud moodustuvad munanditorukestes. Loomulikult on ilma nendeta emase munaraku viljastamise protsess lihtsalt võimatu. Milline on sperma struktuur ja funktsioon?
Sperma avanemine
Spermaraku avastas Hollandi loodusteadlane A. Leeuwenhoek 1677. aastal. Tänu esimestele tema loodud mikroskoopidele suutis teadlane uurida ja visandada mehe sugurakku. Ja ta oli esimene, kes kirjeldas inimese spermat. Seejärel hakkas Leeuwenhoek uurima erinevate loomade isassugurakke. See oli see loodusteadlane, kes esimest korda soovitas neil osaleda eostamise protsessis.
Erinevatel loomadel on isaste sugurakud paigutatud erinevalt, kuigi sperma struktuur on neil väga sarnane:
Pea;
keskmine osa;
Mõnel loomal võib sabavippude arv olla erinev. Erinevat tüüpi elusolendite sperma pea kuju on samuti erinev.
Spermaraku struktuuril, mille foto on ülevaates, on oma iseloomulikud tunnused. Isassugurakkude morfoloogia erineb põhimõtteliselt teistest elusolendite keharakkudest. Lisaks on nende struktuuris kõik peamised organellid. Sperma struktuur on järgmine:
. Pea on ellipsoidne, peaaegu täielikult täidetud tuumaga. See sisaldab isalt järglastele edasi antud pärilikkust. See on esitatud kromatiini kujul, mis on DNA, RNA, valkude kompleks. Tuum sisaldab 23 mehe kromosoomi. See on pool päriliku materjali komplekti. Viljastamise ajal ühineb see emase munaraku 23 kromosoomiga. Spermarakk kannab ühte sugukromosoomi, mille tähis on X või Y. Kui munaraku viljastab Y-kromosoomi kandev meessugurakk, sünnib tulevikus poiss ja kui X on tüdruk. Teisisõnu: sündimata lapse sugu sõltub seemnerakust.

Pea ülaosas (esiosa) asub akrosoom, mis on modifitseeritud Golgi kompleks. See toodab ensüümi hüaluronidaasi, mis on võimeline lagundama mukopolüsahhariididest koosnevat munakoort. Tänu sellele akrosoomi omadusele võib seemnerakk tungida emassugurakku. Pea ja selle kaela piiril on tsentriool - organoid, mis osaleb rakkude jagunemises. See moodustab lipukujulise saba tsütoskeleti. Pea sisaldab ka tsentrosoomi, mis on mikrotuubulite organiseerimise keskus. See tagab sperma saba liikumise, osaleb sügootide tuumade lähenemises ja selle esimeses raku jagunemises.
... Pea ja keskosa eraldav kael... See väike ahenemine sisaldab spiraalset mitokondrit, mis on vajalik energia tootmiseks. See on vajalik sperma aktiivseks liikumiseks. Suurem osa energiast on fruktoosi kujul.
.Vahepealne (keskmine) sektsioon, mida sageli nimetatakse kehaks. See koosneb aksiaalsest keermest. Keskel on mitokondrid, mis sisaldab 28 mitokondrit. Sellel on spiraalne kuju. Mitokondrid sünteesib ATP-d, tagades sellega sugurakkude liikumise.
... Saba on lipukujuline. Selle ristlõikel on näha 9 paari mikroskoopilisi torusid. Veel kaks paari asuvad lipu keskosas. Sperma saba on oluline aktiivse liikumise organoid.

Sperma suurus
Isassuguraku suurus on tühine. Sperma on palju väiksem kui sigoot (välja arvatud saba). Need on teiste inimrakkudega võrreldes väikseimad. Mehe sperma pikkus on umbes 50-70 mikronit, laius 3,5 mikronit. Keskmise osa pikkus on 4,5 µm ja saba 45 µm. Tähelepanuväärne on, et mõnel elusolendil on sugurakkude suurus palju suurem. Nii on näiteks vesiliku sperma pikkus umbes 500 mikronit ja hiire isased sugurakud on 1,5 korda suuremad kui inimese omad. Sugurakkude väiksus on tingitud vajadusest läbida üsna pikki vahemaid munarakuni.
Meeste sperma omadused
Sperma struktuur ja funktsioon on omavahel tihedalt seotud. Isassugurakul on selle eesmärgi ja omaduste tõttu järgmised omadused:
Liikumisvõime tänu nööritaolisele sabale, tänu millele on võimalik kohtuda spermaga ja munarakuga.
Kannab negatiivset elektrilaengut, mis takistab meessugurakke spermas kokkukleepumist.
Terve mehe ejakulaat (seemnevedelik, sperma) sisaldab ligikaudu 200 miljonit spermat. Erinevates elusolendite liikides võib isassugurakkude arv dramaatiliselt erineda. Näiteks hobuse ejakulaat sisaldab ligikaudu 100 miljardit spermat.
Tänu saba-lipule arendab seemnevedelikus olev isassuguraat kiirust kuni 5 cm tunnis.

Isaste sugurakkude omadused
Eksperdid suutsid välja selgitada, et sperma pea tsütoplasmas on vedelkristall. Tänu sellele saavutatakse isase suguraku vastupidavus ebasoodsatele keskkonnatingimustele. Sperma on võimeline vastu pidama naise vagiina agressiivsele happelisele keskkonnale. Spermaraku struktuur muudab selle ioniseeriva kiirguse suhtes vastupidavamaks. Mõnel loomal on isase sugurakul akrosoom, mis on võimeline väljutama muna kinnipüüdmiseks mõeldud pikka niiti.
Sperma elu
Pärast moodustumist munanditorukestes hoitakse spermatosoidid kuu aega munandimanuses. Ejakulaadis püsivad nad elus umbes 24 tundi. Sperma elab tupes umbes 2-2,5 tundi. Kui ta suutis tungida emakakaela või munajuhadesse, pikeneb tema eluiga 2-3 päevani.

Munaraku struktuur
Emaslooma sugurakk on suur, sfääriline, liikumatu rakk, mis on varustatud erinevate toitainetega, mis on vajalikud valkude biosünteesiks ja spetsiifiliste regulatoorsete ainete tootmiseks. Selle munakollane toidab embrüot kogu embrüo perioodi vältel. Selle läbimõõt ulatub 150-170 mikronini.
Munarakku kaitseb väljastpoolt kiirgava krooniga kaetud membraan. Seda ümbritsevad follikulaarse epiteeli rakud, mis küpsedes paljunevad. Nad eritavad spetsiifilist vedelikku. See koguneb primaarsete folliikulite õõnsusesse. Selle epiteeli rakud on seotud munaraku toitainetega varustamiseks. Emassuguraku kest mitte ainult ei toida sugurakku, vaid kaitseb seda ka rohkem kui ühe seemneraku eest. Kuna munarakul ei ole aktiivseks liikumiseks mõeldud aparaati, iseloomustab seda plasma segregatsioon.

Liikumisprotsessi tähtsus
Meeste suguraku liikuvus on selle peamine kvalitatiivne omadus. Seda annab suguraku saba, sooritades sama tüüpi liigutusi. Munaraku ja seemnerakkude struktuursed omadused muudavad viljastamisprotsessi kõige tõenäolisemaks. Isassuguraku kestal on spetsiaalsed retseptorid, mis on võimelised ära tundma munast vabanevaid kemikaale. Tänu sellele võimele on spermatosoidid võimelised pigem eesmärgipäraseks kui kaootiliseks liikumiseks. Pärast ejakulatsiooni suunatakse peaaegu kõik terved meessoost sugurakud emassugurakke. Seda liikumist nimetatakse positiivseks kemotaksseks.

Kõrge spermatosoidide liikuvus mängib olulisemat rolli kui arv ejakulaadis. Seda arutavad sageli meeste ja naiste sugurakkudega töötavad spetsialistid. Seega, kui seemnevedelikus on umbes nelikümmend protsenti spermatosoididest liikuvad, peetakse seda juba patoloogiaks. Sel juhul väheneb oluliselt munaraku viljastamise tõenäosus.
Kui sperma koosneb täiesti liikumatutest spermatosoididest, viitab see sageli sellisele patoloogiale nagu akinospermia. Sel juhul on sugurakud elus, kuid nad on liikumatud ega suuda seetõttu munarakku viljastada. Kõige sagedamini on selle rikkumise põhjuseks sugunäärmete mitmesugused patoloogiad.
Väetamise protsess
Igal meessugurakul on Y- või X-kromosoom, mis määrab munaraku viljastamisel lapse soo. Kõige sagedamini on sellesse protsessi kaasatud üks isassuguraat ja üks emassuguraat. Mõnel juhul viljastatakse rakku 2-3 või enama spermatosoidiga, mis viib ühemunakaksikute sünnini.
Munaraku ja spermatosoidi ehitus on selline, et viljastamise ajal on ainult isassuguraku pea võimeline tungima naise sugurakku. Sel juhul jäävad kõik selle ülejäänud osad väljapoole. Viljastamise (munaraku ja sperma sulandumise) käigus moodustub sügoot, mis on diploidne rakk, mis sisaldab täielikku topeltkromosoomide komplekti.
- See on isase sugurakk, mille põhiülesanne on naise suguraku viljastamine. See sisaldab geneetilist teavet, mis edastatakse isalt sündimata lapsele. Isase suguraku struktuur on täielikult allutatud põhieesmärgile: selle suurus on väike, see on kiire ja mobiilne ning spermatosoidide arv on tohutu. Uurime üksikasjalikumalt, millised nende sugurakkude omadused võimaldavad neil oma funktsioone võimalikult tõhusalt täita.
Meeste sugurakk erineb suuresti mitte ainult emase munarakust, vaid ka kõigist teistest keharakkudest. Kuidas ta ejakulaadis välja näeb, näete ainult mikroskoobi all. Selle suurus on 50-55 mikronit.
Sperma koosneb mitmest osast:
- Pea. Selle kuju näeb välja nagu lusikas. See sisaldab järgmisi põhistruktuure:
- tuum. See sisaldab 23 kromosoomi (22 tavalist ja ühte X või Y, mis määrab sündimata lapse soo). Kui sperma sisaldab X-kromosoomi, on loode emane ja kui Y, siis sünnib poiss;
- membraani vesiikul või akrosoom. Selle struktuuri suurus ja kuju on ligikaudu samad kui südamiku suurus. See sisaldab spetsiaalseid ensüüme, mida sperma vabastab munarakule lähenedes. Nad lahustavad selle membraani ja võimaldavad isassugurakul tungida munaraku tsütoplasmasse;
- tsentrosoom. See struktuur vastutab sabaosa liikumise reguleerimise eest.
- Kael. Pehme osa, mis tagab pea teatud liikuvuse ja võimaluse seda kerge nurga all kallutada. Selle suurus on väga väike.
- Kere või keskosa. Siit läbib telgniit, mis tagab saba liikuvuse. Lisaks sisaldab see mitokondrite kompleksi, mis toodavad energiat, tänu millele saab see rakk liikuda.
- Saba. Koosneb fibrillidest, mis toimivad "kruvina" ja võimaldavad rakul liikuda soovitud suunas. Selle kuju aitab säilitada seemnerakkude normaalset liikumiskiirust ja -suunda.
Huvitav ! On olemas teooria, et meeste sperma (koosY-kromosoom) on aktiivsemad, kuid elavad vaid päeva, kuid X-kromosoomiga on nad vähem liikuvad, kuid vastupidavamad kuni 3-4 päeva. Seetõttu peab poisi eostamiseks vahekord toimuma täpselt õigel ajal.

Küpsemise ja spermatogeneesi tunnused
Isaste sugurakkude küpsemine algab meestel puberteedieas ja kestab kogu elu. Keskmiselt on selle raku arengutsükkel 2,5-3 kuud, seega toimub spermatosoidide uuenemine ligikaudu kord 80-90 päeva jooksul.
Isassugurakud moodustuvad munandites, kus läbivad kõik järjestikused jagunemise etapid. Spermatogeneesi keerulise protsessi käigus moodustuvad spermatiididest küpsed spermatosoidid. Neid spermatogeneesi protsesse reguleerivad munandite ja hüpofüüsi hormoonid.
Mehe organismis on spermatosoidid praktiliselt liikumatud ning ejakulatsiooni (ejakulatsiooni) ajal aktiveeruvad eesnäärme sekretsiooni ensüümid. Ejakulaat sisaldab suurel hulgal neid rakke, umbes 1-2 miljonit spermatosoidi milliliitri kohta.
Tähtis! Tähtis pole spermatosoidide arv, vaid nende liikuvus, normaalsete vormide protsent ja spermatosoidide kontsentratsioon ejakulaadis. Ainult siis, kui need parameetrid vastavad normile, saavad nad oma funktsioone täita.
Kuidas need rakud liiguvad
Kui sperma satub naise tuppe, omandab suur hulk ejakulaadis sisalduvat spermat liikumisvõimet. Neil pole muud funktsiooni, kui liikuda munaraku poole, et seda viljastada.
Tupes on nende kiirus minimaalne, kuid seal toimub esimene spermatosoidide valik ning emaka kehasse jõuavad vaid visad ja liikuvad isendid. Seal suureneb nende kiirus järsult ja nad liiguvad munajuhadesse. Sperma tupest munajuhasse liikumiseks kulub vaid paar tundi ja see on raku mikroskoopilist suurust arvestades väga suur kiirus.

Tupes enamik neist sureb ja liiguvad aeglaselt, sest seal on happeline keskkond. Emakas on keskkond aluseline ja seal nad kiirenevad ja suudavad säilitada oma liikuvuse pikka aega kuni 3-4 päeva.
Emakast suunatakse meessoost sugurakud munajuhadesse, kus sel ajal peab munarakk neid laienenud otsas ootama. Kui seda ei juhtu, liiguvad nad juhuslikult munajuhas mitu päeva ja seejärel surevad. Kogu tupest munarakku liikumise aeg on 1-1,5 tundi, eeldusel, et nende kiirus vastab normile.
Munarakuga kohtumisel on oluline, et spermatosoidide arv oleks vähemalt 300 tuhat. See on vajalik naise suguraku kaitsva membraani lahustamiseks. Kui spermatosoidide kontsentratsioon on madalam, ei pruugi sellest piisata, et "munaraku kaitseriietusest lahti riietada".
Tähtis ! Sperma ellujäämismäära tõttu võib viljastumine toimuda isegi siis, kui vahekord oli paar päeva enne seda.

Küsimused arstile
küsimus: Kui seemneraku ehitus ja kiirus on häiritud, kas see võib meestel põhjustada viljatust?
Vastus: Oh, kindlasti. Kui ejakulaadis on meessoost sugurakkudes struktuursed muutused, on viljastumine peaaegu võimatu. Isegi kui munarakk viljastatakse sellise defektse spermaga, põhjustab see varases staadiumis raseduse katkemise.
küsimus: Kas spermatosoidide arvu ja nende struktuuri saab mõne uuringuga kontrollida?
Vastus: Oh, kindlasti. Tavaline spermogramm näitab spermatosoidide arvu spermas. Lisaks saate läbi viia laiendatud Krugeri spermogrammi, mis võtab arvesse kõiki ebanormaalseid ja liikumatuid vorme.
küsimus: Kas spermat on võimalik ejakulaadis näha palja silmaga või ainult mikroskoobi all?
Vastus: Selle raku suurus on väga väike, seda on ejakulaadis näha ainult mikroskoobi all. Kuid mitte iga seadme all; visualiseerimiseks peate kasutama teatud eraldusvõimega mikroskoopi.
Küsimus: Kui kaua kulub spermatogeneesi parandamiseks?
Vastus: Toitumine, elustiil ja geneetika mõjutavad meeste spermatogeneesi paranemist. Sugurakkude uuenemine toimub iga kolme kuu tagant. Kui olete mures spermatogeneesi parandamise pärast, peate selle aja jooksul välistama infektsioonid ja põletikulised haigused ning seejärel oma elustiili uuesti üles ehitama. Et täpselt teada saada, kui kaua see aega võtab, saate alles pärast kõiki uuringuid.
Meeste suguelundite sugurakkude struktuur on otseselt seotud selle funktsiooniga. Ebanormaalsused nii spermatosoidide kujus kui ka arvus võivad põhjustada probleeme rasestumisel. Diagnoosi selgitamiseks peate tegema spermogrammi ja konsulteerima arstiga.
Sissejuhatus
Inimese sperma
Järeldus
Kirjandus
Sissejuhatus
Spermatozomid (vanakreeka keelest ureEsmb (perekond urEsmbft) - seeme, zhshYu - "elu" ja e? Dpt - "liik") - meessoost sugurakk, isassugurakud, mis viljastavad naissugurakke, mune. Seda terminit kasutatakse väikeste, tavaliselt liikuvate sugurakkude tähistamiseks organismides, mis on oogaamsed. Tavaliselt on need munast palju väiksemad, kuna need ei sisalda nii märkimisväärses koguses tsütoplasmat ja neid toodab organism samaaegselt märkimisväärses koguses. Mõistet "sperma" tuleb eristada mõistest "sperma", kuna viimane koosneb spermast (mis sisaldab spermat) ja sisaldab ka väikest arvu kusiti epiteelirakke. Sünonüümid: elav, mõnikord sperma. Tavaliselt (eriti botaanikas) on spermatosoidid ilma lipukateta.
Sperma mitmekesisus loomadel
Erinevatel loomaliikidel on spermatosoidid erineval viisil paigutatud, kuid siiski on ühiseid struktuurseid tunnuseid. Tüüpilise looma sperma puhul on pea, keskosa ja saba (flagellum) isoleeritud. Pea sisaldab haploidset tuuma (kannab kromosoome), akrosoomi (kannab munaraku membraani lahustamiseks vajalikke lüütilisi ensüüme) ja tsentriooli, mis moodustab lipukese tsütoskeleti. Pea ja keskosa vahel on raku ahenemine, nn kael. Keskel on mitokondrid – hiiglaslik spiraalne mitokondrid. Lipu kasutatakse sperma liikumiseks.
Enamikul loomadel on spermatosoididel ülalkirjeldatud tüüpiline struktuur. Kuid on ka erandeid. Lipude arv võib olla rohkem kui üks. Seega kannavad akvaariumis kalade tetradooni spermatosoidid kahte viburat. Mõnedel vähilaadsetel on spermatosoididel mitu lipukest. Ümarusside puhul on spermatosoididel üldiselt vibud (evolutsiooni käigus on seda tüüpi loomade kõik rakud kaotanud ripsmed ja vibud), neil on amööboidne kuju ja nad liiguvad pseudopoodide abil. Veebil on sabal "lainetav membraan" (uim). Sperma pead on väga mitmekesised. Inimestel on sperma pea munajas, külgedelt lapik. Hiirtel ja rottidel on see konksukujuline. Madalamatel koorikloomadel on sfäärilised spermatosoidid. Mõnel kukkurloomal on spermatosoidid kahekordistunud ja liiguvad paarikaupa, samal ajal sabaga pekstes. Eraldamine toimub vahetult enne munaraku viljastamist.
Spermatosoidid on mikroskoopilise suurusega, reeglina on spermatosoidide pikkus mitukümmend kuni mitusada mikromeetrit. Sperma suurus on samuti väga erinev ega ole korrelatsioonis täiskasvanud looma suurusega. Näiteks hiire sperma on 1,5 korda suurem kui inimese sperma. Ja vesikonna spermatosoidid on mitu korda suuremad kui inimese spermatosoidid.
Inimese sperma
Sperma avanemine
Struktuur ja funktsioon
Inimese sperma on spetsialiseerunud rakk, mille ehitus võimaldab tal täita oma ülesannet: ületada naise sugutrakti ja tungida munarakku, et viia sinna mehe geneetilist materjali. Sperma, ühinedes munarakuga, viljastab selle.
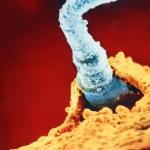
Joonis 1.Inimese sperma struktuur (elektronmikroskoopiline diagramm). 1- akrosoom; 2 - südamik; 3 - kael; 4 - mitokondrid; 5 - aksiaalsed niidid.
Inimese kehas on sperma keha väikseim rakk (kui arvestada ainult pead ennast ilma sabata). Inimese sperma kogupikkus on ligikaudu 55 µm. Pea on umbes 5,0 µm pikk, 3,5 µm lai ja 2,5 µm kõrge, keskmine osa ja saba on vastavalt umbes 4,5 ja 45 µm pikkused.
Väike suurus on tõenäoliselt vajalik spermatosoidide kiireks liikumiseks. Sperma suuruse vähendamiseks selle küpsemise ajal toimuvad erilised transformatsioonid: tuum muutub tihedamaks tänu ainulaadsele kromatiini kondensatsioonimehhanismile (tuumast eemaldatakse histoonid ja DNA seondub protamiini valkudega), suurem osa tsütoplasmast väljutatakse. spermast nn "tsütoplasmaatilise tilga" kujul ainult kõige olulisemad organellid.
Mehe sperma on tüüpilise ehitusega ja koosneb peast, keskosast ja sabast.
Inimese spermatosoidi pea on ellipsoidi kujuga, külgedelt kokku surutud, ühel küljel on väike lohk, seetõttu räägitakse mõnikord inimese sperma pea "lusikakujulisest" kujust. Sperma peas asuvad järgmised rakustruktuurid:
1) Tuum, mis kannab ühte kromosoomikomplekti. Sellist tuuma nimetatakse haploidseks. Pärast sperma ja munaraku (mille tuum on samuti haploidne) ühinemist moodustub sügoot – uus diploidne organism, mis kannab ema ja isa kromosoome. Spermatogeneesi (sperma arengu) käigus moodustuvad kahte tüüpi spermatosoidid: need, mis kannavad X-kromosoomi ja need, mis kannavad Y-kromosoomi. Kui munaraku viljastatakse X-i kandva spermaga, moodustub emasloom. Kui munarakk viljastatakse Y-d kandva spermaga, moodustub isane embrüo. Sperma tuum on palju väiksem kui teiste rakkude tuumad, see on suuresti tingitud sperma kromatiini struktuuri ainulaadsest korraldusest (vt protamiinid). Tugeva kondensatsiooni tõttu on kromatiin inaktiivne – spermatosoidi tuumas ei sünteesita RNA-d.
2) Akrosoom – modifitseeritud lüsosoom – membraanvesiikul, mis kannab lüütilisi ensüüme – aineid, mis lahustavad munakoori. Akrosoom võtab enda alla umbes poole pea mahust ja on ligikaudu võrdne tuumaga. See asub tuuma ees ja katab poole tuumast (seetõttu võrreldakse akrosoomi sageli korgiga). Munarakuga kokkupuutel paiskab akrosoom välja oma ensüümid ja lahustab väikese osa munamembraanist, luues seeläbi väikese "käigu" sperma sisenemiseks. Akrosoom sisaldab umbes 15 lüütilist ensüümi, millest peamine on akrosiin.
3) Tsentrosoom on mikrotuubulite organiseerimise keskus, tagab spermatosoidide saba liikumise, samuti osaleb oletatavalt sügoodi tuumade konvergentsi ja sigooti raku esimese jagunemise protsessis.
Pea taga on seemnerakkude nn "keskmine osa". Keskosa eraldab peast kerge ahenemine - "kael". Saba asub keskosa taga. Lipu tsütoskelett, mis koosneb mikrotuubulitest, läbib kogu sperma keskosa. Lipukese tsütoskeleti ümber paiknevas keskmises osas on mitokondrid – sperma hiiglaslik mitokondrid. Mitokondril on spiraalne kuju ja see on justkui mähitud ümber lipukese tsütoskeleti. Mitokondrid täidab ATP sünteesi funktsiooni ja tagab seeläbi lipu liikumise.
Saba ehk flagellum asub keskosa taga. See on keskmisest õhem ja sellest palju pikem. Saba on spermatosoidide liikumise organ. Selle struktuur on tüüpiline eukarüootse raku viburatele.
Inimese spermatosoidide liikumine
Inimese seemnerakk liigub lipu abil. Liikumise ajal pöörleb sperma tavaliselt ümber oma telje. Inimese spermatosoidide liikumiskiirus võib ulatuda 0,1 mm-ni sekundis. või rohkem kui 30 cm tunnis. Naisel jõuavad esimesed spermatosoidid ligikaudu 1–2 tundi pärast vahekorda koos ejakulatsiooniga munajuha ampullaarsesse ossa (osa, kus toimub viljastumine).
Mehe kehas on seemnerakud passiivses olekus, nende lipuliikumised on tähtsusetud. Spermatosoidide liikumine mööda meeste suguelundeid (seemnetorukesed, munandimanusejuha, vas deferens) toimub passiivselt kanalilihaste peristaltiliste kontraktsioonide ja kanali seinte ripsmete peksmise tõttu. Sperma muutub aktiivseks pärast ejakulatsiooni eesnäärme mahla ensüümide mõju tõttu neile.
Sperma liikumine mööda naise sugutrakti on sõltumatu ja toimub vedeliku liikumise vastu. Viljastumiseks peavad spermatosoidid katma umbes 20 cm pikkuse tee (emakakaela kanal - umbes 2 cm, emakaõõs - umbes 5 cm, munajuha - umbes 12 cm).
Tupekeskkond on spermale kahjulik, sperma neutraliseerib tupehappeid ja pärsib osaliselt naise immuunsüsteemi toimet spermatosoididele. Tupest liiguvad spermatosoidid emakakaela suunas. Sperma liikumissuund määratakse keskkonna pH tajumisega. See liigub happesuse vähenemise suunas; Vagiina pH on umbes 6,0, emakakaela pH on umbes 7,2. Reeglina ei jõua suurem osa spermatosoididest emakakaela ja surevad tupes (postkoitaalses testis kasutatud WHO kriteeriumide järgi ei jää 2 tundi pärast suguühet tuppe elusaid spermatosoide). Emakakaela kanali läbimine on spermatosoididele raskendatud, kuna selles on emakakaela lima. Pärast emakakaela läbimist satuvad spermatosoidid emakasse, mille keskkond on spermatosoididele soodne, emakas suudavad nad oma liikuvust säilitada pikka aega (individuaalsed seemnerakud kuni 3 päeva). Emaka keskkonnal on spermatosoididele aktiveeriv toime, nende liikuvus suureneb oluliselt. Seda nähtust nimetatakse "mahtuvuseks". Edukaks viljastamiseks peab emakasse sisenema vähemalt 10 miljonit spermat. Emakast suunatakse spermatosoidid munajuhadesse, mille suuna ja sees spermatosoidide voolu määrab ära vedelikuvool. On näidatud, et spermatosoididel on negatiivne reotaksis, st kalduvus liikuda ülesvoolu. Vedeliku voolu munajuhas tekitavad epiteeli ripsmed, samuti toru lihaseina peristaltilised kokkutõmbed. Suurem osa spermatosoididest ei jõua munajuha lõppu, nn lehtrisse ehk ampulli, kus toimub viljastumine. Mitmest miljonist emakasse sattunud spermatosoidist jõuab munajuha ampulli vaid paar tuhat. Kuidas inimese spermatosoidid munajuha lehtrist üles leiavad, jääb selgusetuks. On oletatud kemotaksise olemasolu inimese spermatosoidides – liikumist teatud ainete suunas, mida eritavad munarakk või seda ümbritsevad follikulaarsed rakud. Hoolimata asjaolust, et kemotaksis on omane paljude väliselt viljastatud veeorganismide spermatosoididele, ei ole selle esinemist inimeste ja imetajate spermatosoidides veel tõestatud.
Inimese sperma eluiga
Pärast umbes 64-päevast küpsemisperioodi võib mehe sperma säilida kuni kuu aega. Ejakulaadis suudavad nad ellu jääda olenevalt keskkonnatingimustest (valgus, temperatuur, niiskus) kuni 24 tundi. Tupes surevad seemnerakud mõne tunni jooksul. Emakakaelas, emakas ja munajuhas püsivad seemnerakud elus kuni 6 päeva.
Sperma taimeriigis
Enamasti on taimede spermatosoidid väga väikesed; erandiks on tsükaadi spermatosoidid: mõnel liigil on need palja silmaga nähtavad, ulatudes 0,3 mm läbimõõduni. Taimede spermatosoidide tuum on tavaliselt suur, vähese tsütoplasmaga. Taimede spermatosoide nimetatakse ka anterozoadeks. Taimede organit, milles sperma moodustub, nimetatakse anteridiumiks.
Meeste sugurakkude moodustumine
Selgroogsetel moodustuvad naiste sugurakud sugunäärmetes - munasarjades ja isasloomades - munandites. Just sugunäärmetes moodustuvad algsetest diploidsetest rakkudest haploidsed sugurakud. Küpsete spermatosoidide moodustumine imetajate kehas algab puberteediea alguses ja munarakkude moodustumine naisorganismi arengu sünnieelsel perioodil.
Sugurakkude areng
Pesitsusperiood MITOOS
Faasidevaheline kasvuperiood
profaas-I metafaas-I
anafaas-I telofaas-I
Valmimisperiood MEIOOS
profaas-II metafaas-II
anafaas-II telofaas-II
Sugurakkude arengus eristatakse mitmeid etappe (vt diagrammi). Idurakkude arengu esimest etappi nimetatakse paljunemiseks. Seda etappi iseloomustab diploidsete rakkude jagunemine mitoosi teel. Sel juhul moodustub igast emarakust kaks diploidset tütarrakku. Mitoosi tõttu suureneb rakkude arv.
Siis tuleb kasvuetapp. Sel perioodil suureneb rakkude suurus. Rakud on interfaasi seisundis. Nendes sünteesitakse valgud, süsivesikud, lipiidid, ATP, kromosoomid kahekordistuvad.
Küpsemisetapis jagunevad rakud meioosi teel. Kromosoomide arv väheneb poole võrra ja igast diploidsest rakust moodustub neli 1000. haploidset tütarrakku.
Isastel on kõik meioosi tagajärjel tekkinud rakud ühesugused, terviklikud. Sugurakkude areng lõpeb moodustumise perioodiga, mille jooksul moodustuvad sugurakud - sperma ja munarakk.
Sugurakkude teke katteseemnetaimedes toimub omapärasel viisil. Sugurakud moodustuvad tolmukates ja munandites. Tolmukate tolmukad sisaldavad palju diploidseid rakke, millest igaüks jaguneb meioosi teel.Selle tulemusena moodustub igast diploidsest rakust neli haploidset rakku, mis muutuvad õietolmu teradeks See õietolmu moodustumise protsess ei lõpe Iga õietolmutera haploidne tuum jaguneb mitoosi teel Nii moodustuvad kaks haploidset rakku - generatiivne ja vegetatiivne Generatiivne rakk jaguneb taas mitoosi teel, mille tulemusena moodustub kaks haploidset spermat. Sperma – isassugurakud Nad on liikumatud, kuna neil puuduvad lipud ja need viiakse munarakku õietolmutoru kaudu.
Seega sisaldab küps õietolmu tera kolme rakku: vegetatiivset ehk õietolmutoru rakku ja kahte spermat.
Munasarjas on munarakk, milles moodustub naise sugurakk. Munarakus moodustub ühest diploidsest rakust meioosi tulemusena neli haploidset rakku. Kolm rakku surevad ja ülejäänud üks jaguneb mitoosi teel kolm korda. Nii tekib kaheksa haploidset rakku, mis moodustavad embrüonaalse koti. Üks neist muutub munaks, kaks rakku ühinevad ja moodustavad diploidse raku - embrüonaalse koti sekundaarse tuuma. Ülejäänud viis rakku mängivad abistavat rolli, moodustades embrüokoti seina.
Järeldus
sperma meessoost reproduktiivrakk
Sperma ehk sperma, loomade ja paljude taimede küps isasloomarakk. Sperma põhiülesanne on munaraku viljastamine, s.o. isaorganismi geneetilise materjali ülekandmine naise sugurakku. Spermatogenees (sperma moodustumine) toimub enamikul loomadel munandite seemnetorukestes. Selle protsessi kestus erinevatel liikidel ei ole sama: hiirtel on see umbes 35 päeva, inimestel - 74 ja hooajalise paljunemisega loomadel - palju kauem. Imetajatel reguleerib spermatogeneesi (nagu ovulatsiooni) hüpofüüsi folliikuleid stimuleeriv hormoon. Emaslooma suguelunditesse kinni jäänud spermatosoidid võivad püsida elujõulisena pikka aega: mesilastel - 3-4 aastat, lindudel - umbes 3 nädalat; inimestel on nad aga emakas elujõulised mitte kauem kui 48 tundi. Taimedel on spermatosoidid reeglina väga väikesed, enamikus seemnerakkudes on nad liikumatud, lipuvabad, rohe- ja pruunvetikates, sõnajalgades, korte, lükopoodide, tsükaatide, hõlmikpuu ja mõnede teiste taimede puhul aga vastupidi. , spermatosoidid on liikuvad ja neil on kaks või mitu lipukest.
Kirjandus
1. NS Kurbatova, EA Kozlova "Üldbioloogia loengukonspektid".
2. Rusen-Range E. Spermatogenees loomadel. M., 1980.
3. Gilbert S. Arengubioloogia, t. 1. M., 1993.
4. Giljarov MS Bioloogia entsüklopeediline sõnastik. M., 1986.
5. Antonova O. Vanuse anatoomia ja füsioloogia. M., 1995.
Sarnased dokumendid
Sugurakkude küpsemise protsess. Mitmete algloomade, vetikate, eoste, seemneseemnete ja mitmerakuliste loomade elutsükkel. Meeste sugurakkude areng, mis toimub hormoonide regulatiivse mõju all. Spermatogenees inimestel.
esitlus lisatud 01.04.2013
Inimese paljunemise füsioloogilised omadused. Kaks tüüpi sugurakke: isased (sperma) ja emased (munad). Sugurakkude (gameetide) moodustumise protsess on gametogeneesi nähtus. Kolm arenguperioodi: spermatogeneesi, ovogeneesi ja meioosi faasid.
kursusetöö, lisatud 05.04.2009
Meeste suguelundite sisemine struktuur: eesnääre, munandikott ja peenis. Naise sisemiste suguelundite struktuur. Veenid, mis kannavad verd kõhukelmest. Kuulmisorganite funktsioonid. Auditoorne taju inimese arengu protsessis.
abstraktne, lisatud 16.10.2013
Rakumembraani mõiste, ehitus ja funktsioon. Kloroplastide ja mitokondrite struktuur. Lehtede liigid vastavalt lehelaba, serva ja aluse kujule. Võrsete hargnemine ja hargnemine. Keeruliste ja lihtsate õisikute struktuur, odra, rukki, nisu, maisi lilled.
test, lisatud 27.11.2011
Käsnad: struktuur, elupaik, tähtsus looduses ja inimelus. Klassi harilikud ja lubjakäsnad. Bodyagi jõe üldvaade. Koelenteraatide üldised tunnused. Torkavate rakkude põhifunktsioon. Paljunemise ja regenereerimise tunnused.
esitlus lisatud 16.01.2014
Drosophila välisstruktuur ja värvus. Drosophila ontogeneesi kestus ja esimese paaritumise, munaraku ja viljastamise tunnused. Munarakkude ja spermatosoidide küpsemine, soo määramine. Drosophila genoom ja selle kasutamine geneetilises modelleerimises.
esitlus lisatud 26.10.2015
Elusrakkude peamised tüübid ja nende struktuuriomadused. Eukarüootsete ja prokarüootsete rakkude ehituse üldplaan. Taime- ja seenerakkude struktuuri tunnused. Taimede, loomade, seente ja bakterite rakkude ehituse võrdlustabel.
kokkuvõte lisatud 12.01.2016
Transgeneesi meetodid loomakasvatuses. Munandite sugurakkude kasutamine. Transgeenide ekspressiooni suurendavad tegurid loomadel. In vitro kultiveeritud rakkude tuumade siirdamise tunnused. Loomakasvatuse geenitehnoloogia töö väljavaated.
abstraktne, lisatud 26.09.2009
Spermatogeneesi tunnused, mitootiline rakkude jagunemine meioosi tüübi järgi. Rakkude, mis koos moodustavad spermatogeense epiteeli, diferentseerumise etappide uurimine. Meeste suguelundite ja nende näärmete ehituse, eesnäärme funktsioonide uurimine.
abstraktne, lisatud 12.05.2011
Seedesüsteemi näärmed. Naiste suguelundite klassifikatsioon. Meeste suguelundite klassifikatsioon ja struktuur. Südame kambrid ja veresooned, mis toidavad südant. Vere väljavoolu teed. Aju osakonnad. Kuulmis-, nägemis-, liikumiskeskuste lokaliseerimine.
Sperma- keeruka struktuuriga isase sugurakud, millel on iseseisva liikumise omadus, on sperma peamine bioloogiline osa. Sperma koosneb peast, kaelast, kehast ja sabast.
Põllumajandusloomade spermatosoidide kogupikkus on 0,056-0,07 mm ehk 56-70 mikronit ja laius 0,001-0,005 mm ehk 1-5 mikronit.
Sperma maht on väga väike. See on 10-12 tuhat korda väiksem kui muna maht.
Sperma pea on lusikakujuline plaat. Aeglaselt liikuvaid spermatosoide mikroskoobi all uurides saab näha nende pöörlemist ümber pikitelje. Sperma pea pöördub kas laia küljega või servaga, mis paneb sperma “virvema” märgatavalt.
Sperma pea moodustab umbes üheksandiku selle pikkusest. Pärilik teave on koondunud peasse, samal ajal kui kael, keha ja saba on sperma motoorsed aparaadid. Pea eesmine osa on ümardatud ning pulli ja jäära spermatosoidide tagumine osa on mõnevõrra kitsenenud ja järsult ära lõigatud. Pea keskel ja taga on tuum ning ees on spetsiaalne korgikujuline keha - akrosoom. See eritab ensüüme, mis viljastamise käigus lahustavad munaraku kiirgava võra viskoosset ainet, soodustavad sperma tungimist munarakku.
Väljaspool on pea kaetud tiheda läbipaistva kestaga - valgulise aine membraaniga. See katab ka kaela, keha ja saba. Ainult saba otsaosa on kestast vaba. Sperma pikaajalisel säilitamisel membraan paisub ja koorib pallide või tükkidena. Sellised spermatosoidid kaotavad oma võime viljastada. Pea on kehaga ühendatud kaelaga. Kolm õhukest niitide (fibrillide) kimpu, mis moodustavad aksiaalse niidi, ulatuvad pea põhjast kaela. Aksiaalne niit, mis on ümbritsetud kaksikheeliksiga, läbib sperma keha. Sperma keha on sperma vardakujuline osa, mis on peenema saba alus. Saba koosneb aksiaalsest viiest, mis on kaetud troonispiraaliga. Saba otsaosa ei ole kaetud spiraali ja kestaga. See koosneb aksiaalsetest filamentfibrillidest ja näeb välja nagu pintsel.
Sperma liikumine... Spermadel on iseseisev liikumisvõime. Liikumiskeskus asub kaelas ja kehas, seetõttu on pea ärarebimisel spermatosoidide keha liikumisvõimeline ja pea jääb liikumatuks.
Spermas võib täheldada kolme tüüpi liikumist:
- otsekohene kui spermatosoidid liiguvad aktiivselt sirgjooneliselt, on see füsioloogiliselt normaalne liikumine;
- areeni liikumine- pöörlevad ümber pea või liiguvad ringi, selline liikumisvorm viitab sperma kahjustusele, pea tursele, lusikataolise kuju kadumisele;
- võnkuv liikumine- paindub vasakule ja paremale ilma edasi liikumata.
Sperma liikumine toimub saba abil. See paindub ühele küljele ja sirgub seejärel kiiresti. Saba liigutus kordub kiiresti, selle vedelikust tõrjumise tulemusena liigub sperma edasi. 1 sekundi jooksul annab härja sperma saba 37 ° nurga all 9 lööki. Pea lusikakujuline kuju koos saba ühesuunaliste liigutustega viib selle pöörlemiseni piki pikitelge. Sabalöökide kombinatsioon pöörlemisega ümber pikitelje viib spermatosoidide translatsioonilise liikumiseni, spermatosoidide liikumiskiirus on üsna suur - 4-6 mm minutis.
Sperma võnkuvad liikumised tekivad energiaprotsesside aeglustumise tõttu, seda võib täheldada normaalsetes spermatosoidides pärast pikka säilitamist, madalatel temperatuuridel ja keskkonna happelisel reaktsioonil. Sperma kuumutamine, leelistamine põhjustab sirgjoonelise liikumise taastamise.
Normaalsed spermatosoidid liiguvad vedelikuvoolu vastu. Seda liikumise tunnust nimetatakse reotaksiks. Tänu reotaksisele liiguvad nad munajuhas munaraku suunas. Surnud, liikumatud, kantakse vedelikuvooluga kaasa.
Sperma on negatiivse elektrilaenguga. Samanimeline elektrilaeng tagab spermatosoidide vastastikuse tõrjumise, takistades nende kokkukleepumist. Elektrilaengu mõjul asetsevad spermatosoidid paralleelselt, mis loob nende liikumises teatud korra. Elektrilaengu vähenemine nõrgestab spermatosoidide vastastikust tõrjumist, nad hakkavad peade või muude osadega kokku kleepuma. Seda nähtust nimetatakse aglutinatsiooniks.
Sperma aglutinatsioon tekib siis, kui sperma sisaldab positiivselt laetud ioone või spermatoaglutiniine, söötme suurenenud happesus (pH 5-6,4).
Aglutinatsioon võib olla pöörduv, kui sperma rakud kleepuvad kokku ainult peast ja säilitavad saba liikuvuse – "stellaatne aglutinatsioon" ja pöördumatu, kui sperma rakud kleepuvad kokku juhuslikult. On tõestatud spermatoaglutiniinide olemasolu emaste suguelundite piirkonnas võrreldes teise loomaliigi spermaga. Sperma aglutiniinide moodustumine organismi reaktsioonina toimub sperma valgu parenteraalsel tungimisel kehasse, mis toimub sperma korduval ja rohkel sisestamisel emaste suguelunditesse. Noortel emasloomadel spermatoaglutiniinid ei ole, kuid paljupaaritel on. Looduslikes tingimustes sisaldab tervete loomade follikulaarne vedelik ja eesnäärme eritised antiaglutiniinid, mis takistavad aglutinatsiooni ja isegi taastavad sperma normaalse seisundi tähtkujude aglutinatsiooni ajal.
Kui leiate vea, valige tekstiosa ja vajutage Ctrl + Enter.
Sperma moodustumise protsess (spermatogenees) toimub pidevalt ja võib sõltuda tootjate söötmis-, hooldus- ja kasutustingimustest, nende tervislikust seisundist, vanusest ja individuaalsetest omadustest.
Munandimanuse erinevatest osadest pärit spermatosoididel on erinev ellujäämis- ja viljastamisvõime. Parim liikuvus, ellujäämis- ja viljastamisvõime on munandimanuse kanali sabaosas paiknevatel spermatosoididel, kõige halvemini munandimanuse peas paiknevatel spermatosoididel.
Ejakulatsiooni ajal segunevad munandimanuse kanalitest pärit spermatosoidid abisugunäärmete eritistega. Lisasugunäärmete eritised lahjendavad paksu sperma massi ja suurendavad sperma kogumahtu. Pulli puhul lahjendatakse munandimanuse kanalist pärit sperma lisasugunäärmete sekretsiooniga 4–5 korda, jääral 2–3 korda, täkul ja kuldil 20–50 korda.
Spermatosoidide sulandumise tulemusena abisugunäärmete eritistega moodustuvad spermatosoidid. Seega on sperma seerum munandimanuse ja täiendavate sugunäärmete (eesnäärme-, vesikulaar-, Cooperi- ja ureetra) saladus. Sperma, mida mees eritab vahekorra ajal, nimetatakse ejakulaadiks.
Munandimanuse sekreedi ja üksikute abisugunäärmete suhe, nende roll sperma vedela osa (plasma) moodustamisel on spetsiifilised (tabel 8.1).
Tabel 8.1
Sperma plasma komponendid (N.I. Poljantsevi järgi)
Suguelundite ehituse iseärasused ja suguühte dünaamika määravad eri liikide meestel nii ejakulaadi suuruse kui ka koostise variatsioonid. Täkul on ejakulaadi maht keskmiselt 50-100 ml (maksimaalselt 600 ml); metssiga - 200-400 (1000); jääras - 1,0-2,0 (3,5); pull - 4-5 (15); meessoost - 2 (18); kuke jaoks - 0,3 (2,0).
Sõltuvalt looma tüübist on spermatosoidide kontsentratsioon erinev. Vaginaalse seemendusega loomadel on sperma kontsentratsioon 5-10 korda kõrgem kui emaka tüüpi seemendusega loomadel. Niisiis leiavad nad 1 ml spermas keskmiselt: jääras - 2,50-3,50 miljardit, pullis - 0,8-1,2 miljardit, kuldis - 0,15-21 miljardit, täkust - 0, 10-0,15 miljardit sperma .
Keemilise koostise poolest on sperma üks keerulisemaid kehavedelikke. Sperma massist umbes 90–98% moodustab vesi, 2–10% kuivainet, millest umbes 60% valk. Sperma peamised koostisosad on valgud ja lipiidid. Valk sisaldab aminohappeid. Fosforit sisaldav letsitiin on sperma lipiidide hulgas esikohal. 100 ml jäära spermat sisaldab kuni 355 mg fosforit, pulli - 82 mg, kuldi - 66 mg ja täku - 19 mg. Samuti on kaaliumi, naatriumi, kloori, kaltsiumi, magneesiumi, rauda, tsinki ja muid elemente.
Spermas tuvastatakse ensüüme hüaluronidaas, peroksidaas, trüpsiin, antitrüpsiin, amülaas, lipaas jne. Ensüümide kontsentratsioon eri liikide loomade spermas on väga erinev.
Spermas leidub askorbiinhapet, tiamiini (V 1), riboflaviin ( 2), retinool (L) ja teised.Nende arv varieerub olenevalt loomatüübist. Niisiis leiti pullide spermast askorbiinhapet 14 mg%, kuldi - 4 mg%. Teiste vitamiinide sisaldus pullide spermas jääb vahemikku 0,089–0,371 mg%.
Sperma keemilises koostises on olulisel kohal suhkur (fruktoos), mis on spermatosoidide energiaallikas.
Sperma koostis, kogus ja bioloogilised omadused sõltuvad suuresti tootja tingimustest (toitmine, hooldamine, opereerimine jne).
Sperma struktuur. Spermatosoidid koosnevad peast, keskosast ja sabast (joonis 8.1).
Akrosoom
keskosa
Mitokondrid
Riis. 8.1. Sperma struktuuri diagramm (V.S.Shipilovi järgi)
Plasma membraan
Sperma pea -ümar plaat. See sisaldab pärilikku ainet – desoksüribonukleiinhapet, mis annab järglastele üle vanema omadused. Pea on kaetud õhukese tuumaümbrisega, millel on palju poore. Esikülje tuumaümbrise peal on seda kaitstud kork - akrosoom, mis eritab ensüümi hüaluronidaasi, mis on vajalik munarakkude viljastamiseks sperma abil. Vanades spermatosoidides kaob akrosoom ja nad kaotavad võime viljastada. Lõpuks kaetakse kogu pea väljastpoolt kõige õhema sarvkestaga – membraaniga. Kui membraan on kahjustatud, kukub akrosoom maha ja sperma kaotab viljakuse.
Sperma kael sisaldab jämedate fibrillide laiendatud algosi, moodustades eesmise tsentriooli, ja mõnevõrra edasi õhukeste fibrillide esialgseid osi - tagumist tsentriooli.
Proksimaalne tsentriool on ümbritsetud kahe basaalgraanuliga ellipsoidsete rõngaste kujul. Igast basaalgraanulist väljuvad üheksa õhukest fibrilli – fibrillid – ning välimise rõnga fibrillid on mitu korda paksemad kui sisemise rõnga vastavad fibrillid. Fibrillid on omavahel ja aksiaalse niidiga ühendatud õhukeste niitide - kudumisvardadega.
Rõngasfibrillid sisaldavad kontraktiilseid valke (skantiini, spermosiini), mis on koostiselt sarnased aktomüosiiniga. Fibrillide kokkutõmbumisel saba paindub, mõned fibrillid painutavad saba paremale, teised vasakule.
Keha sperma on spiraalkeere, mis keerdub ümber korpuse sees kulgeva teljesuunalise keerme. Keritud filament koosneb ovaalsetest mitokondriaalsetest teradest, mis on üksteisega ühendatud nagu helmed pikaks spiraalselt keerdunud filamendiks (või mitmeks filamendiks). Mitokondrid on eraldatud vaheseintega, millel paiknevad järjestikku oksüdatiivsed ensüümid. Vaheseinad on kastetud rasvakihi sisse. Mitokondrite sees toimub toitainete (peamiselt glükoosi, fruktoosi) oksüdatsioon, energia vabanemine ja oksüdatsiooni käigus erilise aine - adenosiintrifosfaadi (ATP) moodustumine, mis on vajalik sperma saba kokkutõmbumiseks.
Aksiaalne niit koosneb fibrillidest (filamentidest): kaks keskmist, üheksa peenikest ja üheksa jämedat. Õhukesed ja jämedad fibrillid paiknevad kahes kihis ümber keskmiste. Õhukesed fibrillid on kahekordsed, moodustavad sisemise kihi ja nende taga asuvad jämedad fibrillid - välimise. Elektrokeemilised uuringud on näidanud, et jämedad fibrillid sisaldavad adenosiintrifosfataasi, ensüümi, mis lagundab adenosiintrifosfaati ja merevaikhappe dehüdrogenaasi, mis osaleb lagunenud adenosiintrifosfaadi redutseerimises.
Fibrillid on ümbritsetud spiraalsete elementidega. Sperma keha on tihedalt põimunud topeltspiraaliga, mis lõpeb rõngakujulise tsentriooliga, mis asub sperma keha ja saba piiril. Saba kogu pikkuses (välja arvatud otsaosa) on ümbritsetud kolmekordse spiraaliga. Spiraalsed elemendid annavad lipule tugevust ilma paindlikkust kaotamata. Kehapiirkonnas on nad mitokondriaalset päritolu. Mitokondrites on koondunud glükolüütilised ja oksüdatiivsed ensüümid, mis osalevad sperma liikumiseks vajaliku energia tootmises.
Väljaspool on sperma keha kaetud membraaniga. Väävlit sisaldavate aminohapete (tsüstiin, tsüsteiin) sisalduse tõttu selle koostises on see vastupidav proteolüütiliste ensüümide, leeliseliste ja happeliste lahuste suhtes.
Sperma saba koosneb ainult membraaniga kaetud aksiaalsest filamendist, aksiaalfilament koosneb kahest kesksest ja üheksast õhukesest fibrillist. Jämedad fibrillid koos mitokondriaalsete filamentidega lõpevad saba alguses. Telgniit on põimitud peenikese sarvniidiga, mis kaitseb fibrillid lagunemise eest.
Sperma omadused. Seemenduse edukus sõltub spermatosoidide elujõulisusest, vastupanuvõimest väliskeskkonnale ja emaslooma paljunemisaparaadile, nende viljastumises osalemise aktiivsusest ja liikuvusest. Sõltuvalt sperma seisundist ja ümbritsevatest tingimustest jääb liikumise intensiivsus, näiteks jäära sperma, vahemikku 1,9–15,4 mm minutis. On kindlaks tehtud, et pulli seemnerakud suudavad liikuda keskmiselt 4,02 mm 1 minutiga, jäär - 4,6 mm, täkk - 5,22 mm, isasloom - 2,88 mm, kukk - 1,02 mm. .
Vastsaadud spermatosoide mikroskoobi all uurides võib märgata, et üks sperma liigub aktiivselt edasi, teised - ringis, teised ainult kõiguvad, neljandad jäävad liikumatuks ning mida kauem vaatlust tehakse, seda rohkem aega möödub spermatosoidide vastuvõtmise hetkel muutub see nõrgemaks spermatosoidide liikumiseks, seda rohkem spermatosoide ilmub võnkuva liikumisega ja liikumatuna.
Spermade liikumise erinev aktiivsus sõltub nende küpsusest, vanusest, erutuvusest, aga ka välistegurite mõjust neile üldiselt ja eelkõige vedelast keskkonnast, kus nad asuvad. Spetsiaalsed uuringud on kindlaks teinud, et sperma füsioloogiline seisund ei ole sama. Seega on munandimanuse peast saadud spermatosoidid välismõjude suhtes väga ebastabiilsed, võrreldes munandimanuse sabast või ejakulaadist saadud spermaga. Munandimanuse peast pärit spermatosoidid võivad väljaspool keha elada mitu minutit või tunde ning munandimanuse sabast pärit spermatosoidide eluiga arvutatakse päevades. Munandimanuse peast pärit spermatosoidid muutuvad happelisse keskkonda paigutamisel väga kiiresti liikumatuks ja munandimanuse sabast pärit spermatosoidid säilitavad võime liikuda happelises keskkonnas. Küpsete spermatosoidide stabiilsus tuleneb nende lipoproteiinist kattest, mis tekib spermatosoidide liikumisel mööda munandimanuse kanalit. Sperma lipoproteiidkatte säilimine spermaga töötamise ajal kunstliku viljastamise (säilitamine, lahjendamine) ajal on spermatosoidide elujõulisuse seisukohalt väga oluline, kuna kate annab neile sama (negatiivse) elektrilaengu, mis takistab nende aglutineerumist mõju all. elektrijõududest.
Kõigil normaalsetel küpsetel spermatosoididel on sirgjooneline, pealine edasiliikumine. Sperma liigutamine toimub selle saba painutamise ja kiire sirgumisega, vibratsiooni tulemusena tõrjutakse sperma ümbritsevast vedelikust. Saba kokkutõmbed toimuvad samal tasapinnal. Pea kumer-nõgusa pinna tõttu aga pöörleb sperma ümber oma pikitelje. Löökide kombinatsioon sperma pöörlemisega ümber oma telje määrab selle liikumise sirguse. Lisaks tekivad iga sabalöögiga sperma peas vedelikuvoolud ja see libiseb mööda liikuma hakanud vedelikukihte, moodustades "mikrokeerised". Sperma saba läbib neid, võttes ära osa keerisesse kogunenud pöörlemise kineetilisest energiast. Keerised suruvad spermat edasi. Sperma sabade liigutused seerumis suruvad neid edasi ning kõigi spermatosoidide negatiivsed elektrilaengud takistavad nende kokkupõrget. Seetõttu põhjustab paksus spermas kahe jõu koosmõju (spermaste elektrostaatiline tõrjumine üksteisest ja nende aktiivne edasiliikumine) spermatosoidide paiknemises ja liikumises mõningase korrastatuse, mis väljendub keeriselaadsete liigutuste tekkes ejakulaadis. , nähtav palja silmaga. Pööriste liigutuste raskusastme järgi saab hinnata looma sperma kvaliteeti.
Praegu seletatakse spermatosoidide liikumist mööda naise suguelundite piirkonda reotaksega, s.o. omadus liikuda vastu vedelikuvoolu. Reotaksise nähtus spermatosoidides on eksperimentaalselt tõestatud. Avanenud munajuha venitatud pinnale asetatud spermatosoidid liiguvad mööda limaskesta, ületades ripsmete vastupanu ja nende liikumisest tingitud vedelikuvoolu.
Sperma liigub väga intensiivselt. Bishopi ja Gray uuringute kohaselt annab sperma temperatuuril 37 °C üheksa lööki sekundis sabaga, liigutades iga löögiga 8,3 mikronit edasi. Seega läbib sperma igas teises 74,7 mikronit ja minutis 4462 mikronit, see tähendab, et ühe minutiga läbib see vahemaa, mis on 60–70 korda suurem kui selle pikkus. Mõnel loomal liiguvad spermatosoidid veelgi kiiremini.
Sperma aglutinatsioon- spermarakkude liimimine peade või kogu kehaga negatiivse elektrilaengu nõrgenemise või neutraliseerimise tõttu. Sperma aglutinatsioon võib olla pöörduv, kui spermatosoidid kleepuvad kokku ainult peade kaudu ja säilitavad saba liikuvuse - tähtkujuline aglutinatsioon ja pöördumatu, kui spermatosoidid kleepuvad juhuslikult üksteise külge ja on liikumatud (surnud) - spermatosoidide massiivne koagulatsioon.
Sperma elektrilaengute neutraliseerimine toimub sperma happesuse suurenemisel ja mitmevalentsete metallide (kaltsium, magneesium, alumiinium) mõjul. Happesuse (pH) 5,0-6,4 korral toimub tähtede aglutinatsioon. Happesuse edasise suurenemisega suureneb spermatosoidide aglutinatsioon.
On kindlaks tehtud isospermoaglutiniinide olemasolu, mis põhjustavad oma liigi sperma aglutinatsiooni (näiteks lehmade vereseerum võib aglutineerida veise spermat), ja heterospermoaglutiniinide olemasolu, mis toimivad mõne teise liigi looma sperma (näiteks veri). seerum, tupesekretid, emakast väljuv lima põhjustavad teise liigi sperma aglutinatsiooni ega mõjuta oma liigi isaste sugurakke). Sperma aglutiniinide moodustumine on organismi reaktsioon sperma valkude parenteraalsele tungimisele. Seetõttu noortel seemendamiseta emasloomadel spermaglutiniinid puuduvad või on neid vähe ning seemendatud emastel, eriti poeginud emastel, nende arvukus suureneb. Piima või vereseerumit kasutades sperma lahjendina tuleb arvestada sperma aglutinatsiooni võimalusega.
Sperma aglutinatsiooni saab ületada aglutinatsioonivastase reaktsiooniga. Looduslikes tingimustes takistavad naiste suguelundites spermatosoidide aglutinatsiooni munasarja folliikulite vedelikus ja spermatosoidides sisalduvad antiaglutiniinid (eesnäärme sekretsioon). Sel põhjusel on tervete loomade sperma aglutinatsioon haruldane.
Hingamine ja sperma glükolüüs. Sperma liikumine on seotud suure energiahulga kulutamisega. Sperma saavad seda kahe peamise biokeemilise protsessi kaudu: hingamine keskkonnahapniku abil ja fruktolüüs, kui suhkur lagundatakse hapnikuvabas keskkonnas. Kolmas protsess - adenosiintrifosfaadi lagunemine - on abistav. Sperma energia saamiseks kasutatakse keskkonnas või tsütoplasmas leiduvaid aineid. Hingamisel saab sperma oksüdeerimiseks kasutada süsivesikuid, lipiide (fosfatiide) ja valke. Peamiselt oksüdeerivad need aga monosahhariide – fruktoosi ja glükoosi, aga ka glükolüüsi käigus suhkrute lagunemisel tekkivat piimhapet. Kulutades süsivesikuid hingamiseks, saavad spermatosoidid umbes 90% oma energiast liikumiseks.
Sperma oksüdeerib ka mitmehüdroksüülset alkoholi - sorbitooli, mis siseneb sperma vesikulaarsete näärmete sekretsiooni osana. Süsivesikute puudumisel keskkonnas kasutatakse sperma hingamisel lipiide.
Täkkude ja kultide spermas, mis sisaldab vähesel määral suhkrut, kasutavad spermatosoidid hingamiseks peamiselt lipiide. Spermas olevate lipiidide lagunemisel tekib glütseriin, mis hingamise käigus võib oksüdeeruda, moodustades piimhappe, fruktoosi.
Süsivesikute oksüdatsiooni käigus tekib süsihappegaas (CO2), vesi ja suur hulk energiat. Kui üks gramm-molekul suhkruid oksüdeeritakse, tekib 680 000 väikest kalorit energiat.
Hingamisprotsess viiakse läbi spermas sisalduvate ensüümide (tsütokroomoksüdaas, karboksüdaas, dehüdrogenaas jne) abil.
Hingamisprotsessi mõjutavad temperatuur ja keskkonna happesuse või aluselisuse aste. Temperatuuri languse või tõusuga iga 10 ° C kohta hingamine nõrgeneb või kahekordistub. Happelises keskkonnas hingamine nõrgeneb ja leeliselises keskkonnas suureneb. Kvaliteetset spermat iseloomustab kõrge hingamissagedus.
Glükolüüs (fruktolüüs)- energia saamine suhkru lagunemise tõttu hapnikuvabas keskkonnas. See viiakse läbi ensüümide (dehüdrogenaas jne), aga ka fosforhappe osalusel. Glükoosi (fruktoosi) lagunemise tulemusena tekib piimhape ja energia.
Fruktolüüs toodab 20 korda vähem energiat ühe grammi suhkrumolekuli kohta kui hingamine. Seega tarbivad spermatosoidid hapnikuvabas keskkonnas kordades rohkem toitaineid. Kuid see protsess on sperma jaoks oluline, kui sperma siseneb naiste suguelunditesse, kus puudub hapnik.
Oluliselt väljendub fruktolüüs pullide, jäärade ja kitsede spermas, mis sisaldab suhteliselt palju suhkrut. Nende loomade spermat iseloomustab ka kõrge hingamistase.
Suhkrut on täku ja kuldi spermas väga vähe ning fruktolaasi selles sisuliselt ei ole. Nendel loomadel saavad spermatosoidid oma energia peamiselt hingamise kaudu.
Glükoosi või muud suhkrut sisaldava söötmega lahjendatud täku ja kuldi sperma säilib aga pikka aega ilma fruktolüüsi (ehk glükolüüsi) tõttu õhu juurdepääsuta.
Erinevate loomade spermas toimuvate metaboolsete protsesside olemuse järgi jaguneb see kahte tüüpi. Esimesse kuuluvad pulli, jäära, kitse sperma, milles toimub hingamine ja fruktolüüs. Nende loomade sperma on veidi lahjendatud abisugunäärmete eritistega, sisaldab suures koguses suhkrut ja vähe soola ning on intensiivse fruktolüüsiga.
Teine tüüp hõlmab täkkude ja kuldide spermat, milles toimub ainult hingamine. Nende loomade spermas pole peaaegu üldse suhkrut ja fruktolüüsi ei toimu. See on lahjendatud märkimisväärse koguse lisasugunäärmete sekretsiooniga ja sisaldab palju sooli.
Anabioos ja selle tähtsus sperma säilitamisel. Saadud piimhape vabaneb sperma membraani kaudu keskkonda. Piimhappe kogunemine spermasse põhjustab spermatosoidide liikumise aeglustumist või täielikku lakkamist. Sperma langeb samal ajal peatatud animatsiooni olekusse, kus hingamis- ja fruktolüüsiprotsessid on oluliselt pärsitud, mis aitab kaasa sperma pikemale säilimisele.
Sperma läheb ka peatatud animatsiooni olekusse, kui temperatuur langeb. Madalad temperatuurid, aga ka happeline keskkond, pärsivad spermatosoidide eluprotsesse ja samal ajal säilitavad nende energiat. Sperma leelistamine või kuumutamine kehatemperatuurini taastab spermatosoidide liikuvuse.
Seemenduspraktikas kasutatavad sperma säilitamise meetodid põhinevad sperma viimisel peatatud animatsiooni olekusse. Piimhappe suur kontsentratsioon avaldab spermarakkudele kahjulikku mõju.
Sperma liikumine. Peamine erinevus spermatosoidide ja teiste rakkude vahel on nende võime hingamise ja glükolüüsi energia tõttu intensiivselt ja aktiivselt liikuda. Sperma liikumiseks, aga ka lihaste tööks on vaja spetsiaalset ainet – ATP-d. Viljakate pullide sperma sisaldab 7–11 mg%; kui ATP kogus väheneb 3 mg% või rohkem, halveneb spermatosoidide viljastamisvõime. Selgus, et spermas ensüümi spermosiini (lihasevalgu aktomüosiini analoog) mõjul laguneb ATP molekul fosforhappeks ja adenosiindifosfaadiks (ADP). Selle protsessi tulemusena saavad spermatosoidid energiat, mis tagab nende liikuvuse. Spermosiini optimaalset ensümaatilist aktiivsust täheldatakse pH 8,3 juures.
Sperma liikumist on mitut tüüpi - sirgjooneline-translatiivne (sperma liigub aktiivselt sirgjooneliselt edasi), areen (sperma pöörleb ümber pea või liigub ringis, mille raadius on ligikaudu võrdne sperma pikkusega), võnkuv (sperma paindub ühes kohas paremale ja vasakule).
Sperma liikumise keskpunkt asub tema kaelas ja kehas. Kui saba on kehast eraldunud, muutub see liikumatuks, samas kui peata spermarakud saavad edasi liikuda.
Spermatosoidide sirgjooneline-translatsiooniline liikumine on normaalne liikumine ning areen ja võnkumine on patoloogilised. Spermatosoidide liikumine toimub saba abil. See paindub ühele küljele ja sirgub seejärel kiiresti. Sellised saba liigutused korduvad kiiresti ja selle tõrjumise tulemusena vedelikust, milles see asub, liigub sperma edasi. 1 sekundi jooksul annab härja sperma saba temperatuuril 37 ° C üheksa lööki. Sperma pea lusikataoline kuju saba ühesuunaliste liigutuste ajal tagab selle pöörlemise ümber pikitelje. Saba löökide kombinatsioon ümber telje pöörlemisega viib sperma sirgjoonelise-translatsioonilise liikumiseni. Põllumajandusloomade sperma liigub tavatingimustes suurema kiirusega (4-6 mm/min).
Maneeži liikumine ilmneb nõrgestatud spermatosoididel pärast pikka ladustamist või siis, kui pea paisub ja muudab oma tavalist lusikakujulist kuju.
Võnkuvat liikumist täheldatakse siis, kui sperma on nõrgenenud, kui saba liikumine muutub nõrgaks ja aeglaseks. Võnkuvaid liikumisi võib täheldada ka normaalsetes spermatosoidides madalatel temperatuuridel ja keskkonna happelisel reaktsioonil, kui liikumiseks vajaliku energia teke pidurdub. Sperma kuumutamine või leelistamine põhjustab sirgjoonelise translatsioonilise liikumise taastumise.
Normaalsed spermatosoidid liiguvad aeglaselt voolavas voolus ühes suunas – vastu vedelikuvoolu. Seda funktsiooni nimetatakse reotaksseks. Tänu reotaksisele liiguvad spermatosoidid munajuhas munaraku suunas. Surnud, liikumatud spermatosoidid liiguvad koos vedelikuvooluga.
Keskkonnatingimuste mõju spermatosoididele. Spermarakkude elutegevust väljaspool tootja organismi mõjutavad erinevad keskkonnatingimused - valgus, temperatuur, osmootne rõhk sperma vedelas osas, keskkonna reaktsioon, soolalahused, kemikaalid, ravimid, mikroobne ja seenreostus.
Valguse mõju. Hajutatud päevavalgus ei avalda spermarakkudele kahjulikku mõju. Otsese päikesevalguse mõjul spermatosoidide liigutused algul intensiivistuvad ja 20-40 minuti pärast nad surevad. Päikesekiirte negatiivset mõju seletatakse keemilisi protsesse aktiveeriva spektri ultraviolettosa mõjuga spermarakkudele ja infrapunakiirte termilise mõjuga. Seetõttu on seemendamise ajal vaja kaitsta seemnerakke päikese ultraviolettkiirte ja bakteritsiidsete lampide eest.
Kogu sperma käitlemine peaks toimuma siseruumides. Kuna hajutatud valgus seemnerakke ei mõjuta, saab sperma säilitamiseks kasutada tavalisi klaasnõusid või veel parem oranže klaasnõusid, mis püüavad kinni ultraviolettkiired. Seemenduspunktide laboris tuleks akendele riputada nailon- või plastikkardinad.
Temperatuuri mõju spermatosoidide liikumisel ja eluprotsessidel on väga tugev. Loomade kehatemperatuurile lähedasel temperatuuril (37–39 ° C) liiguvad spermatosoidid kõige aktiivsemalt, kuid surevad kiiresti. See on tingitud asjaolust, et sperma aktiivse liikumisega toimub spermas sisalduvate energiaressursside kiire kulumine ning piimhappe märkimisväärne kogunemine (jäära ja pulli spermasse), mis põhjustab seemnete mürgistust. sperma.
Temperatuur üle 39 ° C põhjustab spermatosoidide liikumise kiiret peatumist ja nende surma valgu koagulatsiooni (koagulatsiooni) tõttu.
Kui temperatuur on madalam kui loomade kehatemperatuur, siis spermatosoidide liikumine aeglustub. Sperma järkjärgulise jahutamisega temperatuurini umbes 0 ° C spermatosoidide liikumine peatub ja nad lähevad peatatud animatsiooni olekusse. Kui sperma kuumutatakse temperatuurini 37-39 ° C, taastub nende liikuvus. Madalatel temperatuuridel säilivad spermatosoidid pikka aega elujõulisena. Enamik sperma säilitamise meetodeid hõlmab selle jahutamist. Sperma kiire jahtumine, järsud üleminekud kõrgelt madalale temperatuurile põhjustavad aga spermatosoidides temperatuurišoki. Temperatuurišoki all mõistetakse spermarakkude reaktsiooni kiirele jahutamisele. See väljendub kuumutamisel ja spetsiaalsete lahjenditega lahjendamisel spermatosoidide elustamisvõime kadumises, spermatosoidide inetute vormide ilmumises. Kui sperma jahutatakse kiiresti vähemalt 15-17 ° C-ni, võib spermas tekkida temperatuurišokk. Välja on pakutud palju teooriaid ja hüpoteese, kuid ükski neist ei selgita spermatosoidide temperatuurišoki põhjuseid ja mehhanisme.
F.I. Ostashko usub, et šokimehhanismis mängivad olulist rolli osmootsed ja difusiooniprotsessid. Mida aeglasemalt need protsessid kulgevad, seda vähem avaldub temperatuurišokk. Seetõttu on hiline jahutamine peamine viis spermarakkude temperatuurišoki vältimiseks. Tootjatelt värskelt saadud sperma on kiirele jahutamisele kõige tundlikumad. Temperatuurišokk on eriti lihtne, kui värskelt kogutud sperma jahutatakse alla 18 ° C, seetõttu ei tohiks sperma uurimise ruumide temperatuur olla madalam kui 18 ° C. Pärast värskelt saadud sperma hoidmist toatemperatuuril 1-2 tundi muutub see temperatuuri kiirele langusele vähem tundlikuks. Neutraalse või aluselise reaktsiooniga spermas reageerivad spermatosoidid temperatuurile tugevamini kui kergelt happelise reaktsiooniga spermas.
Madalad temperatuurid mõjutavad elusolendeid negatiivselt, sest külmumisel hävib nende rakkude tsütoplasma (selle kristalliseerumise käigus). Selgitamaks madalate temperatuuride negatiivset mõju spermatosoididele, pakuti välja hüpotees külmutatud tsütoplasma klaaskeha vormi kohta (klaasistumine). Selle hüpoteesi kohaselt võib tsütoplasma jahutamisel tahkestuda klaasjaks (ilma kristalliseerumiseta). Klaaskeha kujul tsütoplasma tahkub ilma vett eraldumata ja ilma molekulide järsu nihkumiseta, mille tulemusena ei häirita selle tavalist struktuuri.
Sperma järkjärgulise jahutamisega temperatuurini 6-10 ° C toimub selles intensiivne kristalliseerumine, mis põhjustab spreemia kiiret surma.
Vedelike klaaskehaliseks tahkestamiseks, kui nende temperatuur läheneb 0 ° C-le, on vaja neid järsult jahutada, et kriitilisest faasist kiiresti läbi libiseda. Tsütoplasma "devitrifikatsiooniks" tuleb spermat kiiresti kuumutada, et toimuks sulamine ja ka ilma kristalliseerumisfaasita.
Arvestada tuleks temperatuurišoki võimalusega, eriti töötamisel veise ja jäära spermaga, milles ejakulaadi väikese mahu tõttu jahtub sperma väga kiiresti. Temperatuurišokk tekib kergesti seemendamisel külmal ajal, külmainstrumentide kasutamisel, kui ei järgita sperma säilitamise ja transportimise reegleid.
Spermatosoidide reaktsiooni temperatuurile tuleks käsitleda kui nende kohanemist evolutsiooniprotsessis. On teada, et munandimanuses on spermatosoidide te-pulgad kehatemperatuurist madalamal temperatuuril loomuliku hõljuva animatsiooni olekus, mille tulemusena suudavad nad oma elujõulisust pikka aega säilitada. Alles pärast spermatosoidide segamist lisasugunäärmete saladusega omandavad nad võime aktiivselt liikuda. Emaslooma reproduktiivseadmesse sattunud spermatosoidid (kehatemperatuuri all) hakkavad liikuma maksimaalse intensiivsusega. See määrab nende kiireima edasiliikumise suguelundite kaudu munajuhadesse ja munaraku suunas.
Sperma lahjendamine söötmega kana munakollasega suurendab spermatosoidide vastupidavust temperatuurišokile.
Et vältida spermatosoidide surma temperatuurišokist, tuleb aretusettevõtetes või seemendusjaamades teha kõik tööd värskelt saadud spermaga temperatuuril 18–25 ° C. Sperma lahjendamiseks kasutatav sööde, tassid, seadmed ja vahendid, mis spermaga kokku puutuvad, peaksid olema sama temperatuuriga.
Kõrge välistemperatuuri kahjulike mõjude vältimiseks (lõunariigid) rajatakse vähesoojusjuhtivast materjalist (puukivi, tellis) aretusettevõtteid ja -keskusi, seemendustöid tehakse varahommikul ja õhtul.
Osmootse rõhu mõju. Nagu teate, tekib kemikaali vees lahustamisel selles osmootne rõhk. Lahuse osmootne rõhk on otseselt võrdeline lahustunud aine kontsentratsiooniga (konstantsel temperatuuril), st. lahustunud ainete kontsentratsiooni suurenemisega vedelikus suureneb ka osmootne rõhk. Lahust, millel on sama osmootne rõhk, nimetatakse isotooniliseks, kõrgsurveks - hüpertooniks, madalrõhuks - hüpotoonseks. Spermas tekitavad osmootse rõhu selles lahustunud soolad, suhkrud ja muud ained. Sperma omakorda võib muuta sperma keemilist koostist, ujutades selle üle oma jääkainetega. Sel põhjusel on spermatosoidide osmootse rõhu väärtus muutuv. Näiteks pullis on 0 ° C juures see vahemikus 4,812–9,142 atm ja keskmiselt 6,95 atm. Sellega seoses on oluline arvestada nii sperma vedela osa füüsikalisi kui ka keemilisi omadusi.
Sperma on väga tundlikud osmootse rõhu muutuste suhtes, st. aine kontsentratsioonile vedelikus, milles need asuvad. Selle osmootne rõhk peaks olema võrdne sperma sisemise osmootse rõhuga. Kui spermatosoidid asetada hüpotoonilisse lahusesse või tavalisse vette, surevad nad kiiresti siserõhu suurenemise tõttu. Hüpotoonilise lahuse mõjul spermatosoidide sabad paisuvad ja keerduvad ringi või poolrõngana. Kui sperma segada hüpertoonilise lahusega, surevad ka spermatosoidid, kuid dehüdratsiooni tõttu. Nad kortsuvad, nende sabad omandavad siksakilise kuju. Osmootse rõhu kiire muutus on spermarakkudele äärmiselt kahjulik. Muudel soodsatel tingimustel võivad spermatosoidid säilitada oma elujõulisuse ainult isotoonilistes lahustes.
Erinevate liikide loomade spermas ei ole osmootne rõhk sama. Niisiis on viinamarjasuhkru lahus jäära sperma suhtes isotooniline kontsentratsiooniga 6,4, pulli ja kuldi - 6, täku - 7,2; küülik - 5,4; koerad - 5-7%. Roosuhkru lahus (sahharoos) on täkkude sperma suhtes isotooniline kontsentratsioonis 11%. Sperma reageerib vähem osmootse rõhu kõrvalekaldele hüpertensiooni suunas kui hüpotensiooni suunas.
Loomade seemendamisel tuleb rangelt jälgida, et kõik sperma lahjendamiseks kasutatavad lahused oleksid isotoonilised ja et sperma ei puutuks kokku veega. Tavaline joogivesi võib sisaldada (olenevalt kohalikest tingimustest) märkimisväärses koguses erinevaid sooli, mistõttu vee soolasisaldust arvestamata valmistatud lahus võib spermarakkudele kahjulikku mõju avaldada.
Sperma on vajalik kaitsta selle vedela osa aurustumise eest, näiteks sperma mahutis kuivamise eest, mikroskoobiga uurimisel ja muudel manipulatsioonidel, mille käigus võib soolade kontsentratsioon spermas suureneda. On vaja kaitsta spermat soolade puudumise eest. Näiteks paar tilka destilleeritud vett, mis on tekkinud seemneraku anuma seintele temperatuurierinevuse tõttu (kondensatsioon), võib avaldada kahjulikku mõju jäära või pulli ejakulaadile.
Keskkonna reaktsiooni mõju. Sperma liikuvus ja ellujäämine sõltuvad nende keskkonna reaktsioonist või selle happesuse astmest (pH). Destilleeritud vesi on neutraalne - pH umbes 7,0; happelise reaktsiooni korral on pH alla 7,0 ja leeliselise reaktsiooni korral üle 7,0.
Mannusmanuses on spermatosoidid happelises keskkonnas (pH 5,57-6,90). Ejakulatsiooni ajal seguneb munandimanuse sisu kõrvalsugunäärmete aluselise sekretsiooniga, mis põhjustab spermatosoidide reaktsiooni neutraalsele või nõrgalt aluselisele (pH 7,0). Värskelt saadud veise ja jäära sperma reaktsioon on nõrgalt happeline (pH 6,5-6,7), kuldi ja täku sperma on aga kergelt aluseline (pH 7,2-7,6), mis on seotud munandimanuse vedeliku suurema või väiksema lahjendusega lisasugunäärmete sekretsioonid.
Värskelt saadud neutraalse või kergelt happelise reaktsiooniga spermatosoidid on elujõulisemad kui aluselise reaktsiooniga spermatosoidid. Säilitamise ajal suureneb piimhappe kuhjumise tõttu pulli ja jäära sperma happesus. Samal ajal on spermarakkude liikumine pärsitud ja need langevad peatatud animatsiooni.
Aluseline keskkond aktiveerib spermatosoidide liikuvust. Täku ja kuldi spermas, mis on lahjendatud suurel hulgal abisugunäärmete aluselise eritisega, tarbivad seemnerakud aktiivses liikumises kiiresti energiaaineid ja elavad suhteliselt lühikest aega. Nende tootjate sperma säilitamisel muutub selle reaktsioon leeliselisemaks. See tekib hingamise käigus tekkiva süsihappegaasi lendumise ja ammoniaagi akumuleerumise tagajärjel, mis suurendab leeliselisust.
Muutused keskkonna reaktsioonis mõjutavad ebasoodsalt sperma elujõulisust. Kuid spermal on võime neutraliseerida drastilisi muutusi oma keskkonna reaktsioonis. Seda sperma omadust nimetatakse puhverdamiseks. Spermapuhvrid on nõrkade hapete (süsinik-, sidrun-, piim-, fosforhape) ja valkude soolad.
Tugeva happe mõjul loovutavad nõrkade hapete soolad sellele happele oma metalli ja neutraliseerivad selle. Leeliste neutraliseerimine toimub samal viisil. Sperma valgud, olles nii nõrgad happed kui ka nõrgad alused, on võimelised siduma happelisi ja aluselisi aineid. Sidrunhappe soolad on sperma tugevaim puhver. Veiste ja jäära spermat iseloomustab kõrgeim puhverdusvõime.
Sperma ellujäämismäär väljaspool keha sõltub happe omadustest. Orgaaniliste hapete (piim-, äädik-, merevaik-, võihape jne) mõjul peatub spermatosoidide liikumine keskkonna kõrgeima happesuse juures. Anorgaanilised happed (НС1, Н 2 80 4, Н 3 Р0 4 ja teised), vastupidi, isegi märkimisväärsetes kontsentratsioonides ei avalda see spermatosoididele negatiivset mõju. Erinevus orgaaniliste ja anorgaaniliste hapete mõjus spermatosoididele on seletatav vabade ioonide võimetusega rakku tungida ja sellest tulenevalt väliskeskkonna H-ioonide võimatusega mõjutada rakusiseseid reaktsioone. Mineraalhapped on lahustes täielikult dissotsieerunud, mistõttu nad ei saa mõjutada spermas toimuvaid reaktsioone. Orgaaniliste hapete lahused sisaldavad alati molekule, mis pole ioonideks lagunenud, need tungivad läbi sperma membraani, lagunevad ioonideks, põhjustades rakusisest hapestumist.
Kemikaalide mõju. Vaatamata kõigi aseptika reeglite järgimisele on saadud spermatosoidid alati enam-vähem saastunud bakteriaalse flooraga. Edu ei krooninud desinfitseerimisvahendi otsimist, mis võiks avaldada bakteritsiidset toimet mikrofloorale, ilma et see spermat negatiivselt mõjutaks. Uuringud on näidanud, et ühtegi kaasaegset desinfektsioonivahendit ei saa pidada spermale kahjutuks. Väikestes annustes, mida pole märgata isegi kõige peenemate keemiliste reaktsioonide käigus, on neil kahjulik mõju spermarakkudele. See seletab loomade kunstliku viljastamise ebaõnnestumist, kui seda tehakse veterinaarhaiglates, ambulatoorsetes kliinikutes või kui kasutatakse samu vahendeid, mida kasutatakse ravi ajal.
Leiti, et seemnerakud tapavad kiiresti elavhõbekloriidi annuses 0,00001 g, kaaliumpermanganaadis annuses 0,00004 g, lüsooli, kreoliin, äädikhappe annuses 0,0003 1 g sperma kohta. Leelised ja happed, eeter, ammoniaak, tärpentin on sperma tugevad mürgid. Ka plii, vase, raua, hõbeda oksiidid on spermale väga mürgised, seetõttu kasutatakse seemendamisel klaasist või nikeldatud instrumente.
Plasmas ja spermatosoidides endas leiduvatel elektrolüütide ioonidel võib olla spermatosoididele mitmekülgne toime: membraani pingutamine või lõdvendamine; neutraliseerida elektrilaeng; muuta membraanide läbilaskvust; aeglustada või vastupidi aktiveerida ainevahetusprotsesse.
Elektrolüüdid koosnevad katioonidest ja anioonidest. Mono- ja kahevalentsed katioonid ei muuda oluliselt sperma ellujäämist. Anioonidel on tugevam toime kui katioonidel ja anioonide toime sõltub nende valentsusest. Kloriidianioonid vabastavad sperma membraani ja hävitavad lipoproteiini membraani. Fosfaatide, sulfaatide, tsitraatide anioonid, vastupidi, paksendavad sperma membraani ja stabiliseerivad elektripotentsiaali. Sellega seoses kasutatakse sperma lahjendamiseks mõeldud söötmete koostises fosfor-, väävel- ja sidrunhappe sooli.
Lenduvate orgaaniliste ainete (lüsool, kreoliin, tärpentin, formaliin, ammoniaak, eeter, jodoform, kseroform) aurud avaldavad spermale kahjulikku mõju isegi eemalt. Sellega seoses on kunstliku viljastamise ettevõtetes ja keskustes keemiliseks desinfitseerimiseks kasutatavate ainete loetelu väga piiratud. Arvesse tuleks võtta ka tubakasuitsu, odekolonni aurude, parfüümi, küüslaugu ja sibula kahjulikku mõju spermale.
Antimikroobsetest ainetest on nitrofuraanid (furatsiliin, furasolidoon) vähetoksilised. Mõned sulfoonamiidid (streptotsiid) ja enamik antibiootikume madalas kontsentratsioonis on kahjutud, seetõttu kasutatakse neid sperma lahjendites.
Hoolimata asjaolust, et põhimõtteliselt on seemenduse läbiviimisel desinfitseerimisvahendite kasutamine täiesti vastunäidustatud, on nad siiski sunnitud kasutama nakkushaiguste ennetamiseks. Selleks on kõige vastuvõetavam puhas rektifitseeritud alkohol. Võrreldes teiste desinfitseerimisvahenditega on seda lihtsam instrumentidelt eemaldada (aurustub kiiresti) ja lahustub hästi vees. Siiski tuleb meeles pidada, et 0,5% alkoholilahus tapab sperma väga kiiresti. Tooralkohol, denatureeritud, ebapiisavalt puhastatud fuselõlidest ja muudest lisanditest, on loomade seemendamiseks täiesti sobimatu.
Mikroobse ja seente saastumise mõju. Loomade mikroobse ja seentega saastatuse astme, sperma kvaliteedi ja viljakuse vahel on otsene seos. Looduslikes tingimustes ei leidu tervetel tootjatel mikroorganisme munandi keerdunud tuubulites, lisandites, seemnejuhades ja kusiti algosas. Seega nakatuvad spermatosoidid urogenitaalkanali läbimisel ja kokkupuutel väliskeskkonnaga. Spermas olevate mikroorganismide kvantitatiivne ja kvalitatiivne koostis varieerub suuresti sõltuvalt tervislikust seisundist, tootja hügieenilisest seisundist, kunsttupe, mänguaeda, labori, sperma säilitusajast ja paljudest muudest teguritest. Selgus, et aretusettevõtete ja seemenduspunktide ruumide õhu uurimisel tuvastati 15 seeneliiki, mille hulgas oli patogeenseid ja mürgiseid. Eriti palju mikroorganisme leidub siseõhus, kus tootjaid hoitakse. Halbade hügieenitingimuste korral sisaldab 1 m 3 õhku kuni 1,6 miljonit mikroorganismi. 1 ml halbades hügieenitingimustes kasvatatud pullide spermat sisaldab 85–230 miljonit mikroorganismi. Kultide puhul on spermatosoidide tõsise mikroobse saastumise allikaks eesnaha divertikulaar (pimekott). Nagu on kindlaks teinud I.G. Moroz ja M.P. Ryazansky, pärast divertikulektoomiat väheneb järsult spermatosoidide mikroobne saastumine, suureneb emiste viljakus ja viljakus.
Sperma saastumine patogeensete mikroorganismidega võib põhjustada seemendamise käigus trihhomonoosi, vibrioosi, brutselloosi, tuberkuloosi, leptospiroosi, suu- ja sõrataudi ning teiste nakkus- ja invasiivsete haiguste levikut. Sperma on hea kasvulava paljude patogeensete ja mittepatogeensete mikroorganismide säilimiseks ja paljunemiseks. Samal ajal ei vähenda ei antibiootikumid ega sperma pikaajalisel säilitamisel madal temperatuur selles mikroobikehade arvu ega taga selle desinfitseerimist, kuna mikroobide liigiline koostis on ebatavaliselt mitmekesine. Spermas elav mikrofloora jaguneb liikide lõikes nelja rühma: saprofüütsed, patogeensed, oportunistlikud, algloomad ja riketsiad.
Tinglikult patogeensest mikrofloorast leidub pullitootjate spermas Pseudomonas aeruginosa ja Escherichia coli, Staphylococcus, Streptococcus, Proteus jt.Pseudomonas aeruginosa on kõige patogeensem.
G.V. Zverev ja A.N. Repko leidis, et värskes ja lahjendatud spermas leidub koos bakteritega patogeensete ja toksiliste omadustega seeni. Nad tuvastasid 21 seeneliiki. Patogeensed seened nagu Candida albicans, Aspergillus fumigatus, Lichtheimia eirymbifera, aitavad kaasa endometriidi tekkele lehmadel ning tiinuse alguses põhjustavad embrüote ja loodete surma.
Sperma sisse kinni jäänud mikroobid paiknevad peamiselt spermatosoidide läheduses ja osa neist on otse spermatosoididel – peas, kaelal, kehal, sabal. Mikroobidega kokkupuutel tekib turse ja seejärel pea perforatsiooni (korgi) ja kogu sperma kesta hävimine, lagunemine.
Seemendamiseks on patogeense mikrofloora ja Pseudomonas aeruginosa puudumisel lubatud sperma, mille mikroobikehade sisaldus ei ületa 5 tuhat. Tootja värske lahjendamata spermatosoidi kolitisaator (väikseim kogus testitavat materjali, mis sisaldab ühte E. coli) ei tohiks olla suurem kui 1:10.
Mineviku kordamine
- 1. Sperma bioloogilise struktuuri ja füsioloogilise seisundi tunnused.
- 2. Spermiogenees. Epididüümi bioloogiline roll.
- 3. Sperma liikumise tunnused mehe suguelundites.
- 4. Väliskeskkonna mõju spermatosoidide ellujäämisele.
- 5. Fruktolüüsi tähtsus pullide, jäärade ja kitsede spermas.
- 6. Spermatosoidide ehituse eripärad erinevatel loomaliikidel.