Морфологія бактерій Що таке морфологія мікроорганізмів
Глава 1. МОРФОЛОГІЯ І КЛАСИФІКАЦІЯ МІКРООРГАНІЗМІВ
Морфологіямікроорганізмів вивчає форму і особливості будови клітин, здатність рухатися, утворювати суперечки, способи розмноження та ін. . Організми, які мають клітинної будови, становлять третє надцарство - акаріоти (наприклад, віруси). До прокаріот відноситься тільки одне царство - бактерії, в тому числі ціанобактерії (синьо-зелені водорості). До еукаріот відносяться три царства: тварини, рослини і гриби.
Мал. 1. Форми бактерій:
а- кулясті; б - паличкоподібні; в- звивисті; 4- нитчасті; д- нові форми- 1 - мікрококи; 2-стрептококи; 3 -диплококи і тетракоки; 4 - стафілококи; 5-сардини; б-палички без суперечок; 7 – палички зі спорами; 8"-вібріони; 9 - спірили; 10 - спірохети; //- Тороїди; 12 - Бактерії, що утворюють простіки; 13 - червоподібні"; 14 - шестикутні
Розподіл живих організмів на прокаріоти та еукаріоти базується насамперед на особливостях будови їхнього ядерного апарату. За допомогою електронного мікроскопа було встановлено відсутність у бактерій істинного ядра, тому їх назвали прокаріотами, тобто доядерними організмами. Відомо, що основою ядерного апарату є дезоксирибонук-леїнова кислота (ДНК), молекула якої має вигляд подвійної спірально закрученої нитки. Ядерний апарат прокаріотів включає молекулу ДНК у вигляді замкнутої в кільце нитки, розташовану безпосередньо в цитоплазмі. Ядерний апарат прокаріотів називають нуклеоїдом, що в перекладі з латинського означає «подібний до ядра». У еукаріотів є справжнє ядро з ядерцем, оточене ядерною мембраною. Усередині ядра укладено ДНК. Поруч із основним ознакою є багато специфічних особливостей у будові та обміні речовин прокаріотів.
Основними об'єктами технічної мікробіології є бактерії, міцеліальні гриби та дріжджі, які в основному складають як корисну, так і небажану мікрофлору харчових виробництв.
ПРОКАРІОТИ (БАКТЕРІЇ)
У світі мікроорганізмів бактерії за чисельністю (близько 4000 видів) та різноманітністю здійснюваних ними хімічних перетворень займають провідне місце. Більшість бактерій це одноклітинні організми, але є і багатоклітинні.
Форма та розміри бактерій. Одноклітинні бактерії на вигляд поділяються на три основні групи: кулясті, паличкоподібні та звивисті (рис. 1).
Кулясті бактерії-коки (рис. \,а)можуть бути одиночними - мікрококиабо з'єднаними попарно - диплококи.Часто при розподілі клітини з тих чи інших причин не розходяться і утворюють різні поєднання, які залежать від розташування перегородки, що ділить. Коли ділять перегородки розташовуються у двох взаємно перпендикулярних площинах, то утворюються групи, що складаються з чотирьох клітин. тетракоки.При розподілі в трьох взаємно перпендикулярних площинах утворюються пакетоподібні скупчення, що складаються з восьми - шістнадцяти коків. сарцинами.При розподілі коків у різних напрямках утворюються скупчення клітин, що нагадують грона винограду, - стафілококи.Якщо поділ коків відбувається в одному напрямку і вони при цьому не поділяються, то утворюються ланцюжки клітин- стрептококи.Ці поєднання не еквівалентні багатоклітинним мікроорганізмам, тому що кожна клітина в них є окремим організмом, здатним на самостійне існування після відокремлення від інших клітин.
 |

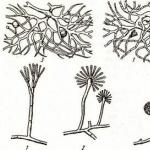
Мал. 2. Актиноміцети:
а- міцелій; б - спороносні
Паличкоподібні бактерії (рис. 1, б)мають форму витягнутого циліндра, можуть бути одиночними або попарно з'єднаними, а також у вигляді ланцюжків з трьох і більше клітин. Відношення довжини клітини до її діаметра у них сильно варіює. У коротких паличок довжина лише перевищує поперечний переріз і їх іноді досить важко відрізнити від коків. Паличкоподібні бактерії є найчисленнішою групою серед бактерій.
Звивисті (рис. 1, в)бактерії бувають трьох типів: вібріони- палички, вигнуті у вигляді коми; спірили,мають кілька правильних завитків, та спірохети,мають вигляд дрібних спіралей з численними завитками.
Крім цих найбільш поширених у природі форм бактерій є невелика кількість нитчастих форм (рис. 1, г).Вони являють собою багатоклітинні організми у вигляді ниток, що складаються з однакових циліндричних або дископодібних клітин.
Порівняно нещодавно в ґрунті та водоймах були виявлені нові форми бактерій, клітини яких мають вигляд розімкнутого або замкнутого кільця (тороїди), шестикутної зірки, розетки, а також клітини з виростами (просте-ками) та червоподібної форми (рис. 1, д).
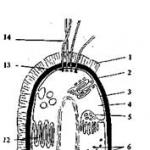
Мал. 3. Схема будови бактеріальної клітини: 1 - капсула; 2 - Клітинна стінка; 3 - цитоплазматична мембрана; 4 - цитоплазма; 5 - мезосоми; 6 - рибосоми; 7 - полісахаридні гранули; 8 - нуклеоід; 9 - включення сірки; 10 - жирові краплі; 11 - Поліфосфатні гранули; 12 - внутрішньоплазматичні мембранні утворення; 13 - базальне тільце; 14 - джгутики
До бактерій належить ще одна, особлива група мікроорганізмів – актиноміцети. Їхні клітини в основному мають вигляд дуже тонких довгих прямих ниток, що гілкуються (рис. 2).
Розміри бактерій мізерно малі, поперечний переріз клітин більшості бактерій вбирається у 0,5-0,8 мкм, середня довжина паличкоподібних бактерій від 0,5 до 3 мкм. Нитчасті бактерії значно більші - не-
які мають 15-125 мкм у довжину і 5-35 мкм у поперечнику. Довжина клітин спірохет може досягати 500 мкм. Найбільш дрібні з мікроорганізмів – мікоплазми, що не мають клітинної стінки, мають розмір 0,1-0,15 мкм.
Об'єм бактеріальної клітини в середньому становить 0,07 мкм 3 маса - 5-10~ 12 г. В 1 мм 3 може міститися до 10 9 бактеріальних клітин.
У харчових виробництвах основне значення мають кулясті та паличкоподібні бактерії.
Будова, хімічний склад та функції клітинних структур бактеріальної (прокаріотної) клітини. Обов'язковими клітинними структурами у більшості бактерій є: клітинна стінка, цитоплазматична мембрана (ЦП*М), ядерний апарат (нуклеоїд) і рибосоми (рис.3).
Клітина зовні покрита жорсткою клітинною стінкою.Вона надає форму клітині, оберігає її від несприятливих зовнішніх теплових та механічних впливів, захищає клітину від проникнення до неї надлишку води. У деяких бактерій на зовнішній поверхні клітинної стінки утворюються капсулиабо слизовий шар.Капсула найчастіше складається з полісахаридів (декстрану, левану), рідше - з поліпеп-тидів. Капсула – необов'язкова структура бактеріальної клітини. Іноді капсули служать джерелом запасних поживних речовин. Наприклад, капсули з полісахаридів утворюються у клітин лейконостоку на середовищах із значною кількістю вуглеводів.
За хімічним складом та будовою клітинної стінки бактерії діляться на 2 великі групи: грампозитивніі грамнегативні бактерії(Трам+ та Грам -).

гіс. *. ч.лема будовиклітинних стінок грампозитивних та грамнегативних бактерій
Названо так на ім'я датського вченого Крістіана Грама, який запропонував спеціальний спосіб забарвлення бактерій (забарвлення за Грамом). Після забарвлення препарат бактерій обробляють спиртом або ацетоном, в результаті чого грам - бактерії знебарвлюються, а грам + бактерії зберігають темно-фіолетове забарвлення. Забарвлення Грама має важливе значення для класифікації бактерій.
Як у Грам + , так і у Грам - бактерій жорсткість
- клітинної стінки обумовленанаявністю полімерного з'єднання
неня пептидоглікану(Муреї-на), але у Грам + бактерій його кількість значно більша (до 90-95% від речовин клітинної стінки), а у Грам - - 5-10%. Пептидоглікановий шар у Грам+ бактерій щільно прилягає до ЦПМ (рис. 4).
Крім того, в клітинних стінках Грам+ бактерій є інші полімери - тейхоєві кислоти,які так само, як і пептидоглікан, є тільки у прокаріотів, а у еукаріотів не виявляються. У складі клітинної стінки Грам+ бактерій у невеликих кількостях містяться полісахариди.У Грам+ бактерій клітинна стінка має товщину 20-80 нм, юна одношарова та щільна.
Клітинна стінка Грам – бактерій значно тонша – 10-13 нм, але вона багатошарова. Пептидоглікан утворює лише внутрішній шар, що нещільно прилягає до ЦПМ. До внутрішнього шару прилягає зовнішня мембрана, що складається з ліпопротеїдіві ліпополісахаридів.Тейхові кислоти в клітинній стінці Грам – бактерій відсутні.
Зовнішня мембрана Грам - бактерій перешкоджає проникненню у клітину токсичних речовин, тому Грам - бактерії значно стійкіше проти Грам+ бактеріями до дії антибіотиків, отруйних хімічних та інших. речовин. Тому в харчових виробництвах боротьба з Грам – бактеріями за допомогою дезінфікуючих засобів не завжди ефективна.
Цитоплазматична мембрана(ЦПМ) розташована під клітинною стінкою, обмежує вміст клітини та відіграє дуже важливу роль у житті клітини. Порушення її цілісності призводить до загибелі клітини. Хімічно ЦПМ є білково-ліпідним комплексом, що складається з білків (50- 75% від маси ЦПМ), ліпідів (в основному фосфоліпіди - 15-45%) і невеликої кількості вуглеводів. У ЦПМ є пори, якими у клітину надходять поживні речовини і виводяться кінцеві продукти обміну речовин.
Оскільки у прокаріотів ЦПМ єдина, на відміну від еукаріотів, мембранна структура в клітині, то вона виконує багато функцій: здійснює транспорт поживних речовин із зовнішнього середовища всередину клітини за допомогою специфічних білків - переносників; на внутрішній стороні ЦПМ розташовані окислювально-відновні ферменти, що беруть участь у постачанні клітини енергією, та гідролітичні ферменти, що здійснюють розщеплення високомолекулярних сполук. У деяких бактерій ЦПМ утворює вп'ячування всередину клітини. мезосоми,що мають різні форми та розміри та виконують різні функції (участь в енергетичних процесах, у процесах поділу клітини, процесі розмноження та ін.).
Цитоплазма- це внутрішній вміст клітини, оточений ЦПМ, що є напіврідкою колоїдною системою. Вона містить воду до 70-80% від маси клітини, ферменти, амінокислоти, набір РНК, субстрати та продукти обміну речовин клітини. У цитоплазмі розташовуються інші життєво важливі структури клітини - нуклеоїд, рибосоми, і навіть запасні речовини різної природи.
Нуклеоїдє ядерний апарат прокаріотів. Це компактне утворення, що займає центральну область в цитоплазмі, що складається з подвійної спірально закрученої нитки ДНК, замкненої в кільце, яка ще називається бактеріальною хромосомою. Бактеріальна хромосома в одній точці стикається з мезосомою. У розгорнутому вигляді нитка ДНК може мати довжину понад 1 мм, тобто майже в 1000 разів більша за довжину бактеріальної клітини. Вся генетична інформація у прокаріотів, так само як і у еукаріотів, міститься в ДНК, тому функція нуклеоїда полягає в передачі спадкових властивостей. Перед поділом клітини нуклеоїд ділиться навпіл. Ядерний апарат прокаріотів не має ядерця і не відокремлений від цитоплазми ядерною мембраною, як це має місце у еукаріотів.
Рибосоми- невеликі гранули, розсіяні в цитоплазмі, що складаються з РНК (60%) та білка (40%). Вони грають дуже важливу фізіологічну роль, оскільки у них відбувається синтез білків. У молодих клітинах спостерігається підвищений вміст рибосом.
У клітинах бактерій, крім обов'язкових клітинних структур, є включення запасних речовин.Вони накопичуються при надлишку тих чи інших поживних речовин у середовищі, а витрачаються при голодуванні клітини. До запасних речовин клітини бактерій відносяться. полісахариди,включають глікоген, крохмаль і гранульоз; жирові краплі,містять ліпіди (жири) у вигляді полі-р-оксимасляної кислоти, яка синтезується на середовищах багатих на вуглеводи. Полі-р-оксимасляна кислота зустрічається тільки у прокаріотів і її кількість може досягати 50% від сухої маси клітин. Гранульози та ліпіди служать хорошим джерелом вуглецю та енергії для клітини. У багатьох прокаріотів у клітинах накопичуються поліфосфати у вигляді гранул, званих також валютиновими або метахроматиновими зернами. Вони використовують клітинами як джерело фосфору.

Мал. 5. Схема прикріплення
1 - Клітинна стінка; 2 - цито-
плазматична мембрана; 3 -
мембрана джгутиків; 4 -диски
основи; 5 - джгутики
У клітинах деяких бактерій, що у перетвореннях сірки, відкладається молекулярна сірка як особливих включень.
Рухливість бактерій.Здібно-
стю до руху має приблизно Уб частина бактерій. Це в основному багато паличкоподібних і всіх звивистих форм бактерій. Нерухомими є майже всі кулясті бактерії (коки), понад 50% паличкоподібних бактерій та ряд інших.
Найчастіше рух здійснюється за допомогою джгутиків(див. рис. 3) тонких ниток товщиною 10-20 нм, що складаються з особливого білка флагеліну.Довжина джгутиків багато разів може перевищувати довжину клітини. Джгутики (рис. 5.) прикріплюються до мембрани за допомогою двох пар дисків основиі через пори в ній та клітинній стінці виходять назовні. Швидкість переміщення бактерій здопомогою джгутиків висока (20-60 мкм/с).
Характер розташування джгутиків лежить на поверхні клітини одна із ознак класифікації бактерій (рис. 6). Їх кількість може бути від 1 до 100. Бактерії, що мають один джгутик на кінці клітини, називають монотрихами;з пучком джгутиків на одному або обох кінцях клітини - лофотріхами;один джгутик на обох донцях - амфітріхами.Бактерії, у яких джгутики покривають всю поверхню клітини, називаються перитрихами.Джгутики забезпечують активний рух клітин лише у рідкому середовищі, і за втрати джгутиків при старінні чи механічному впливі клітини втрачають здатність до руху, але зберігають життєздатність.
До рухомих форм відносяться також спірохети, деякі нитчасті (багатоклітинні) та інші бактерії, що не мають. джгутиків. Спірохети можуть пересуватися і в рідкому середовищі, і по твердому субстрату внаслідок хвилеподібних скорочень клітини. Нитчасті бактерії, ціанобактерії та інші мають ковзний тип руху по твердому і напівтвердому субстрату.
Здатність до руху дозволяє бактеріям переміститися в ту область середовища, в якій умови для їх зростання та розмноження (концентрація поживних речовин та кисню в середовищі, освітленість та ін) найбільш оптимальні.
Мал. 6. Розташування джгутиків у рухомих форм бактерій: а- монотрих; б – амфітріх; в- лофотріх; г -перитріх
Зростання та розмноження бактерій.Основною відмінністю живих організмів від неживої природи є зростання і розмноження. Зріст- це фізіологічний процес, у ході якого збільшуються розміри та маса клітини. Зростання бактеріальної клітини обмежене, і, досягнувши певної величини, вона перестає зростати. Починається процес розмноження,тобто збільшення числа особин (клітин), коли від материнської клітини відокремлюється дочірня.
Більшість бактерій розмножується простим розподілом на дві частини. Такий спосіб розмноження називається бінарним поперечним поділом.У переважної більшості Грам+ бактерій клітини діляться рівно навпіл за допомогою септи(Поперечної перегородки). На протилежних сторонах внутрішньої частини клітинної стінки утворюються два виступи, що ростуть назустріч один одному (від периферії до центру), у цих місцях ЦПМ утворює мезосоми (вп'ячування). Розташовані у мезосомах ферменти синтезують матеріал клітинної стінки. Поперечна перегородка спочатку фомірується з ЦПМ і пептидоглікану; зовнішні шари синтезуються пізніше.
Клітини більшості Грам – бактерій діляться шляхом утворення перетяжки. У центрі клітини з одного боку ЦПМ іклітинна стінка поступово прогинається до злиття з протилежною поверхнею клітини. Утворенню поперечної перегородки або перетяжки передує поділ ДНК, внаслідок чого кожну дочірню клітину потрапляє по одному нуклеоїду.
Актиноміцети розмножуються головним чином екзоспора-ми(зовнішніми спорами), які утворюються поодинці або ланцюжками на кінцях спороносних гіф- спороносцях, мають найрізноманітнішу форму (див. рис. 2). Існують інші способи розмноження.
Освіта ендоспор.Здібністю до освіти ендоспор(Внутрішніх спор) мають лише деякі паличкоподібні Грам + бактерії. Оскільки в кожній клітині утворюється тільки одна суперечка, то спороутворення є не спо-
 |
 |
Мал. 7. Типи спороутворення у бактерій:
а- Бацилярний; б- клостридіальний; в - плектридіальний
собом розмноження, а стадією клітини, що лежить, для перенесення несприятливих умов. Суперечки утворюються при голодуванні, при надлишку продуктів обміну речовин чи невідповідності температури, вологості та рН їх оптимальним значенням у розвиток даного виду бактерій.
Розрізняють три типи спороутворення (рис. 7). Якщо при утворенні суперечки у центрі клітини форма її не змінюється, то такий тип спороутворення називається бацилярним;він властивий представникам роду ВасШіз. Якщо ж клітина в середині потовщується і набуває вигляду веретена, то такий тип спороутворення називається клостридіальним.Іноді суперечка утворюється ближче до кінця клітини і тоді клітина набуває вигляду тенісної ракетки - такий тип спороутворення називається плектридіальним(Мал. 7). Клостридіальний та плектридіальний типи спороутворення властиві бактеріям роду С1оз1:пс1шт.
Спороутворення - складний процес, в результаті якого у клітині формується ендоспора, що відрізняється від вегетативної клітини структурою та хімічним складом (рис. 8). Ендоспора має зовнішню та внутрішню мембрани, між якими розташовується кортекс(Кора), подібний за хімічним складом з клітинною стінкою вегетативної клітини. Поверх зовнішньої мембрани утворюються багатошарові покриви суперечки, які у основному з білків. У деяких бактерій зовні суперечки формується ще один шар. екзоспоріум,що складається з ліпідів та білків.
При спороутворенні відбувається накопичення специфічної речовини – дипіколінової кислоти, яка відсутня у вегетативної клітині, а також іонів кальцію. Процес утворення суперечки протікає кілька годин. Коли суперечка сформується, оболонка та інші частини клітини руйнуються і суперечки звільняється.
Мал. 8. Схема будови бактеріальної суперечки:
/ - Нуклеоїд; 2 - цитоплазма; 3 - Внутрішня мембрана; 4 - кортекс; 5 - Зовнішня мембрана; 6 - покриви, що складаються з кількох шарів; 7 – екзоспоріум
Спори надзвичайно стійкі до впливу температури, наприклад, суперечки збудника важкого харчового отруєння - ботулізму - витримують нагрівання до 100 °С протягом 5-6 год. Спори виносять висушування, вплив ультрафіолетових променів, отруйних речовин тощо. , що їх покриви важкопроникні, в них міститься багато ліпідів, а також дипіколінової кислоти і кальцію. Активність ферментів у них пригнічена. Висока термостійкість суперечка обумовлюється низьким вмістом у них води, що оберігає білки від денатурації при високих температурах.
Суперечки бактерій можуть зберігати життєздатність десятки і навіть сотні років. Потрапивши у сприятливі умови, суперечка поглинає воду і набухає, її термостійкість знижується, зростає активність ферментів, під дією яких розчиняються оболонки, і суперечка проростає у вегетативну клітину.
Псування харчових продуктів викликають лише вегетативні клітини бактерій. Тому необхідно знати умови, що сприяють утворенню спор та їх проростання у вегетативні клітини, щоб правильно вибрати спосіб обробки харчових продуктів з метою запобігання їх псуванню під впливом бактерій.
Принципи класифікації бактерійВ даний час типової класифікації бактерій немає, хоча робота з її створення ведеться постійно. Класифікація всіх живих істот заснована майже повністю на безпосередньо спостерігаються і легко визначуваних морфологічних ознаках організмів. У бактерій внаслідок нечисленності їх морфологічних ознак створити загальноприйняту класифікацію неможливо і потрібні додаткові ознаки.
Крім того, організми відповідно до основних принципів класифікації живих істот повинні бути розташовані в ряди від найпростіших до більш складних, тобто як йшов їх поступовий розвиток (еволюція). Така класифікація організмів є природною.Найдрібнішою одиницею класифікації є вигляд- Група організмів, наділена загальними стабільними ознаками і походить від загального предка. Близькоспоріднені види групуються у вищу систематичну одиницю - рід;близькі пологи - у сімейства,сімейства - у порядкиабо загони,порядки - у класи,а класи - у типи.
Однак мікробіологи в даний час не мають достатніх знань про еволюцію бактерій. Тому більшість наявних класифікацій бактерій є штучними.Штучні класифікації призначаються для визначення тієї чи іншої групи мікроорганізмів, яка представляє практичний інтерес для дослідника.
Наукові назви мікроорганізмів складаються з двох латинських слів: перше пишеться з великої літери і означає рід, друге - з малої літери і означає вид цього роду. Наприклад: ВасШіз зіМШз (палочка сінна) - це бактерія, що відноситься до роду ВасШіз, паличкоподібної форми, що утворює ендоспори бацилярного типу, постійно мешкає на сіні.
Для класифікації бактерій використовують здебільшого такі ознаки: морфологічні(форма клітин, наявність та характер розташування джгутиків, спосіб розмноження, забарвлення за Грамом, наявність ендоспор); фізіологічні(ставлення до впливу температури, рН, кисню, тип харчування, спосіб отримання енергії, характер продуктів, що утворюються); культуральні(Характер зростання на різних поживних середовищах культури бактерій у масі, а не у вигляді окремих клітин: на рідких середовищах це наявність плівки, каламуті, осаду; на щільних середовищах - тип колоній та їх особливості).
В останні роки здобула визнання класифікація бактерій, запропонована Р. Мюрреєм в 1978 р. Це штучна класифікація, в основу якої покладено будову клітинної стінки. Усі бактерії, котрим характерна будова клітинної стінки типу Грам+ бактерій, віднесено до відділу Тчгтаси1.ез*. Інший відділ - ОгасШсі1ез - об'єднує всі бактерії, які мають клітинну стінку, характерну для Грам - бактерій. Третій відділ поєднує спеціальні форми мікробів, позбавлені реальної клітинної стінки; вони не відіграють ролі в харчових виробництвах і тому не розглядатимуться. Бактерії, що мають значення в харчових виробництвах, належать до перших двох відділів.
Відділ р1гтасі1ез. До нього відносяться 4 групи; в основу поділу на групи покладено форму клітин та здатність утворювати ендоспори та екзоспори. Це коки, дві групи паличкоподібних бактерій, актиноміцети та споріднені організми.
Коки характеризуються округлою формою; розподіл клітин відбувається в одній або декількох площинах, при цьому утворюються різні поєднання клітин; коки нерухомі, не утворюють ендоспор. Багато мікрококів є збудниками псування харчових продуктів, лейконосток є шкідником у цукровому виробництві; деякі стафілококи, розвиваючись у харчових продуктах, виробляють отруйні ве-
* Від лат. «кутикула» – шкірка, «фірма» – солідна, «гратія» – витончена.
ства (токсини) і викликають харчові отруєння. Сюди ж відносяться молочнокислі стрептококи, що використовуються при виробництві кисломолочних продуктів, маргарину, вершкового масла та ін.
Друга група – це палички, що утворюють ендоспори. До них належить одне сімейство, представники якого дуже поширені у природі. Це одиночні илц з'єднані в ланцюжки палички, багато з них рухливі, мають перитрихіальне джгутикування. Палички утворюють ендоспори бацилярного типу (рід ВасШиз) і клостридіального або плектридіального типу (рід СЛозхгМшт). Багато хто є збудниками псування харчових продуктів (наприклад, гнильні, маслянокислі бактерії). Є багато збудників інфекційних хвороб (сибірки, правця) і харчового отруєння - ботулізму.
Третя група – це палички, які не утворюють ендоспор. До них відноситься тільки одне сімейство, що включає рід 'ас1о'асШіз. Це паличкоподібні, що не утворюють суперечки молочнокислі бактерії*. Найчастіше вони є одиночними довгими і тонкими паличками, іноді - короткими паличками ланцюжками. Є шкідниками у процесах бродильних виробництв. Використовуються у виробництві кисломолочних продуктів, в. сироділлі, квашенні овочів, у хлібопеченні.
Четверта група - актиноміцети та споріднені організми. Актиноміцети - своєрідна група бактерій, що являють собою довгі тонкі нитки, що гілкуються без перегородок, звані гіфами,переплетення яких утворює міцелій. Нижня частина міцелію, що вростає в субстрат, називається субстратним міцелієм і служить для забезпечення організму харчуванням, верхня частина міцелію підноситься над субстратом і називається повітряним міцелієм.Розмножуються актиноміцети екзоспорами, що утворюються у спороносцях. Деякі з актиноміцетів являють собою короткі палички, що гілкуються. Зустрічаються на харчових продуктах, можуть викликати їх псування, коли продукти набувають виразний землистий запах. Є й патогенні види (туберкульозна та дифтерійна палички). Актиноміцети є основними продуцентами антибіотиків, одержуваних у промислових масштабах, а також вітамінів групи В (В 'В 2, В 3, В 6, В1 2).
Відділ (Зг асШсихез. Всі представники Грам^ бактерій не утворюють суперечку і різко різняться по здатності розвиватися на світлі і без нього. Бактерії, що зустрічаються в харчових виробництвах, байдужі до світла. Вони різняться за формою клітин і способу руху. За кількістю предста-
* Незважаючи на те, що представники цього роду - палички, що не утворюють суперечку, у науковій літературі за ними зберігається стара назва 'ас1о-'асшиз.
 |
вителів іЗначимості в природі та житті людини найбільший інтерес з них представляють псевдомонади та ентеробактерії.
З псевдомонад для харчових виробництв найбільше значення має рід Рзеиоотопаз. Це поодинокі рухливі палички з одним або з пучком полярних джгутиків (монотрихи та лофотріхи). Псевдомонади дуже поширені вприроді, які беруть активну участь у кругообігу речовин, часто виявляються у водоймах і грунті, забруднених різними сполуками, наприклад пестицидами, беруть участь і в їх розкладанні. Багато псевдомонад утворюють флюоресцирующие пігменти, що виділяються в середу, і викликають псування харчових продуктів (деякі гнильні, жиро-окисляючі та інші бактерії).
До Грам - паличок відносяться також оцтовокислі бактерії пологів Асе1оЪае(ег (перитрихи) і 01ісопоЬас1ег (моно-трихи), що використовуються у виробництві оцту, багато з них є шкідниками в бродильних виробництвах.
У харчових виробництвах найбільше значення має численна кишкова група бактерій – ентеробактерії. Це поодинокі рухливі палички, перитрихи, але трапляються і нерухомі форми. Деякі з них постійно населяють кишечник людини та тварин (наприклад, кишкова паличка), інші є збудниками інфекційних шлунково-кишкових захворювань (дизентерії, черевного тифу, паратифу), що передаються через харчові продукти, а також збудниками харчових отруєнь.
Класифікація бактерій, що мають значення в харчових виробництвах і розглядаються в даному курсі, наведено на с. 20.
ЕУКАРІОТИ (міцеліальні гриби та дріжджі)
Одним із трьох царств, що належать до надцарства, еукаріотів, є гриби. Раніше вважали, що гриби займають проміжне положення між царствами рослин і тварин, так як ряд ознак зближує їх як з тваринами, так і з рослинами. Але нині гриби виділено окреме царство Мусо1а. Ця велика і різноманітна група організмів включає до 100 тис. видів.
Гриби поширені у природі. Вони мешкають у різних кліматичних зонах від тропіків до Арктики, особливо багато їх у ґрунтах, у тому числі високогірних, на рослинах; зустрічаються вони у прісних та солоних водоймах, у місцях з підвищеною вологістю тощо. п. Гриби для свого розвитку потребують органічних речовин.
Серед грибів зустрічаються організми, що розвиваються за рахунок органічних речовин відмерлих організмів; вони беруть участь у кругообігу речовин у природі. Але є й такі,

Мал. 9. Міцелій грибів:
а- несептований; б - септований
які можуть існувати тільки в живих організмах та викликати їх захворювання. Деякі з грибів виділяють отруйні речовини. мікотоксини.Багато грибів викликають псування харчових продуктів і пошкодження різноманітних виробів і матеріалів, деякі можуть розвиватися навіть на оптичних поверхнях, де є мізерна кількість мастила. Вони утилізують мастило і викликають помутніння лінз. Але гриби мають і важливе практичне значення, багато хто з них вживається в їжу, використовуються у виробництві етилового спирту, органічних кислот, ферментів, антибіотиків, вітамінів, деяких сортів сиру тощо.
Міцеліальні гриби. Царство грибів ділиться на сім класів, але об'єктами вивчення мікробіології є в основному три, що включають міцеліальні гриби, - зигоміцети (раніше їх називали плісняві гриби), аскоміцети та дейтеро-
Форма та розміри. Клітини міцеліальних грибів мають витягнуту форму у вигляді ниток (гіфів), розміри яких досягають до 5-30 мкм у діаметрі, що значно перевищує розміри бактеріальної клітини.
Переплетення гіф утворює тіло гриба. міцелій,або грибницю(Рис. 9). Більшість гіф розвивається над поверхнею субстрату (повітряний міцелій), де розташовуються органи розмноження, а частина - у товщі субстрату (субстратний міцелій). Гіфи у більшості міцеліальних грибів багатоклітинні, в їх клітинах є поперечні перегородки. септи.Такий міцелій називають септованим, він є у аскоміцетів і дейтероміцетів. Міцелій зигоміцетів несептований і є однією гігантською клітиною з кількома ядрами. Гіфи ростуть за рахунок верхівкових клітин, і клітини гіф неоднакові по довжині.
Деякі гриби на певній стадії розвитку утворюють плодові тіла,всередині яких знаходяться органи роз-

Мал. 10. Схема будови грибної
1 - Клітинна стінка; 2 - ядро; 3 - ядерна мембрана; 4 - рибосоми; 5 - апарат Гольджі; 6 - цитоплазматична мембра-»на; 7-лізосоми; 8 - ендоплазматична мережа; 9 - мітохондрії; 10 - цитоплазма
множення, вкриті зверху щільним переплетенням гіф. В інших видів грибів із щільних переплетень сильно розгалужених гіф утворюються склероцш,багаті на запасні поживні речовини. Вони служать для перенесення несприятливих умов і є формою гриба, що лежить.
Міцеліальні гриби не мають джгутиків і належать до нерухомих організмів.
Будова клітини. У міцеліальних грибів клітини
мають будову, характерну
для клітин еукаріотних мікроорганізмів (рис. 10). У них є добре розвинена система внутрішньоклітинних елементарних біологічних мембран (на відміну від прокаріотів, у яких тільки одна мембранна структура всередині клітин - цитоплазматична мембрана). Внутрішньоклітинні структури еу-каріотів, повністю "обмежені від цитоплазми такими мембранами, називаються органелами.До органелів крім ЦПМ. відносяться ядро, мітохондрії, ендоплазматична мережа, апарат Гольджі та лізосоми.
Зовні клітина міцеліальних грибів покрита багатошаровою жорсткою клітинною стінкою, що складається на 80-90% полісахаридів. Основним із них є азотомісткий полісахарид хітин. Полісахариди пов'язані з білками, ліпідами, поліфосфатами. Під клітинною стінкою розташована ЦПМ, що оточує цитоплазму. У цитоплазмі розташовано ядро;воно містить ядерце, хромосомита оточене ядерною мембраною з порами. У ядерці синтезуються і накопичуються попередники рибосом, які потім транспортуються через пори ядра до цитоплазми. У грибів у длітках буває від одного до 20-30 ядер. У цитоплазмі розпорошені рибосоми.
Мітохондрії- мембранні структури, які грають дуже значної ролі. Вони є багатокамерними мішечками або трубочками з еластичними стінками, що утворюють вп'ячування. кристи(Рис. 11). На них знаходяться окислювально- 
Мал. 11. Схема будови мітохондрій:
а- загальна схема будови; б-подовжній розріз мітохондрії; / - Зовнішня міто-хондріальна мембрана; 2 - Внутрішня мітохондріальна мембрана; 3 - Кристи; 4 - матрикс
відновлювальні ферменти (у прокаріотів ці ферменти локалізовані в ЦПМ), що беруть участь в енергетичному обміні. Тому мітохондрії називають «силовими станціями клітини», «енергетичними ансамблями» тощо.
Ендоплазматична мережа-мембранна система, що складається з канальців, бульбашок або цистерн, які не мають строго певної локалізації, а розташовуються або по периферії клітини, або навколо ядра, або пронизують всю цито
плазму. Там розташовані різні ферменти, відповідальні за синтез ліпідів, вуглеводів, за транспорт речовин усередині клітини. Апарат Гольджі- система мембран, пов'язаних з ядерною мембраною та мембранами ендоплазматичної мережі. Він розташований на ділянці цитоплазми, де немає рибосом. Роль апарату Гольджі повністю не з'ясовано. Припускають, що в апараті Гольджі відбувається синтез матеріалу клітинної стінки та нових мембран, а також з його допомогою здійснюється транспортування речовин, що синтезуються в ендоплаз* матичної мережі, та видалення з клітини продуктів обміну.
Лізосомиявляють собою мембранні структури округлої форми. У них зосереджені гідролітичні ферменти (у прокаріотів вони локалізовані в ЦПМ), що розщеплюють білки, полісахариди, ліпіди.
У клітинах міцеліальних грибів добре видно вакуолі- порожнини, оточені мембраною та заповнені клітинним соком. Зазвичай вони розташовані поблизу клітинної стінки, їх кількість збільшується в міру старіння клітин. Основні запасні поживні речовини міцеліальних грибів – глікоген, який утворюється на середовищах із надлишком цукру; метахро-матин, який у вигляді гранул знаходиться у самих вакуолях, а в цитоплазмі поблизу вакуолей накопичуються ліпіди у вигляді жирових крапель.
Розмноження та класифікація. Міцеліальні гриби розмножуються безстатевим та статевим шляхом. Обидва способи розмноження пов'язані з утворенням спор - зовнішніх (екзо-спори) та внутрішніх (ендоспори). Утворенню спор при статевому розмноженні передує процес злиття вмісту двох клітин та їх ядер. Знову ядро, що утворилося, ділиться на кілька частин - суперечка. Крім того, всі гриби

Мал. 12. Зигоміцети:
1 - КЬ12оріз; б - мисог - спорангій з ендоспорами; в -послідовні стадії утворення зигоспори при статевому розмноженні; г- проросла зігоспора зі спорангієм
можуть розмножуватися вегетативно – шляхом верхівкового зростання
гіф, а також за допомогою шматочків гіф та міцелію. Гриби, спо
особливі до статевого розмноження, відносяться до досконалим
(аскоміцети, зигоміцети), а ті, які не мають статевого
розмноження, відносяться до недосконалимгрибам (дейтеро-
міцети). У грибів є велика різноманітність способів
та органів розмноження. \
Клас 2у^отусе1;е5 (зигоміцети). Це найпростіше організовані гриби. Міцелій у них несептований, багатоядерний, має вигляд однієї гігантської розгалуженої клітини. До зигоміцетів відносяться мукорові гриби. Вони широко поширені у природі. Найбільше значення мають лпредставники пологів Місог та КИгориз.
Розмножуються зигоміцети безстатевим і статевим шляхом (рис. 12). При безстатевому розмноженні в особливих кулястих здуттях - спорангіях,формуються на кінцях довгих плодоносних гіф - спорангієносців,утворюються ендоспори, звані спорангіоспорами.Спорангієносці бувають одиночними (у грибів роду Місог) або зібраними в пучки з коренеподібними розростаннями біля основи - різоїдами (угрибів роду Кигориз).
При статевому розмноженні спочатку відбувається злиття двох багатоядерних гіф міцелію, які є зазвичай короткі утворення з невеликим потовщенням на кінцях. Потім відбувається попарне злиття ядер. Закінчується статеве розмноження освітою зиготи(зігоспори), яка після періоду спокою проростає та утворює коротку гіфу зі спорангієм на кінці. При проростанні суперечки відбувається розподіл ядер. Багатоядерна цитоплазма спорангія розпадається на безліч спорангіоспор, які в сприятливих умовах можуть прорости в міцелій.

Мал. 13. Конідієносці аскоміцетів: а -у грибів роду АзрегдШіз; б- у грибів роду РегпсПшт; / - вегетативний міцелій; 2 - конідіє-носець; 3 - фіаліди; 4 - конідії
Багато грибів роду* Місог викликають псування харчових продуктів утворюючи пухнасті сірі нальоти. Гриби роду КИгориз викликають так звану «м'яку гниль» ягід, плодів та овочів. Борошнові гриби утворюють органічні кислоти і ферменти, здатні викликати слабке спір 1 -тове бродіння, у зв'язку з чим вони застосовуються в деяких країнах Сходу для отримання напоїв.
Клас Аз сот у-се1;е5 (а з комі це-ти, або сумчасті гриби). До них відносяться представники широко поширених грибів пологів РешсШшгп і Аз-рег^Шіз.
Аскоміцети мають добре розвинений багатоклітинний міцелій. Безстатеве розмноження у них відбувається за допомогою зкзоспор, званих конідіями,які формуються на кінцях спеціалізованих ГІФ - конідієносців.У аспергілів вони прості, без перегородок, здуті на вершині у вигляді міхура, на якому розташовуються фіаліди,ланцюжки, що відчленюють, кулястих конідій. У пеніцилів конідіоносці багатоклітинні, у вигляді пензлика, що складається з мутовок фіалід (рис. 13). Конідії бувають різного забарвлення (зелені, жовті, чорні, блакитні тощо). Конідії поширюються повітряними потоками, комахами, краплями роси, дощу та, проростаючи, утворюють новий міцелій.
Статеве розмноження аскоміцетів відбувається шляхом злиття вмісту та ядер двох клітин різних гіф, після чого відбувається розподіл ядра; навколо нових ядер концентрується цитоплазма та утворюється спорова оболонка. Материнська клітина покривається товстою оболонкою і перетворюється на аск(сумку), усередині якого найчастіше знаходиться 8 аскоспор. Зверху сумка покривається переплетенням гіф, утворюючи плодове тіло.

Мал. 14. Конідіоносці та конідії різних пологів недосконалих грибів: а- Во1гуІз; б- різагшт; в -АІєгпану; г -С1ас1о5ропит
Однак деякі представники сумчастих грибів знайшли практичне застосування. Так, окремі представники пеніциллових грибів використовуються як продуценти антибіотика пеніциліну в промислових масштабах, інші – у виробництві сиру сортів «Рокфор», «Камамбер». Аспергили продукують органічні кислоти, у зв'язку з чим застосовуються для промислового отримання лимонної кислоти (Азрег§Шиз ш§ег). Багато аспергіл використовуються для промислового отримання різних ферментних препаратів, що використовуються в галузях харчової та легкої промисловості.
Клас Оеі1еготусе1ез (дейтероміцети). Дейтероміцети, або недосконалі гриби, мають багатоклітинний міцелій. Статеве розмноження у них відсутнє, вони розмножуються тільки безстатевим шляхом, в основному конідіями, які, як і конідіоносці, мають різну форму і вигляд.
Конідіоносці найчастіше багатоклітинні, але можуть бути одиночні - розгалужені або у вигляді пучків, зі здуттями. Конідії можуть бути одноклітинними, багатоклітинними, іноді з поздовжніми та поперечними перегородками (рис. 14). За формою конідії бувають кулясті, еліпсоподібні, нитки-
 |
 |
видні, серповидні, зірчасті та ін. Деякі дейтеро-міцети (наприклад, молочна пліснява) розмножуються не конідіями, а особливими клітинами - артроспорами,які утворюються в результаті фрагментації конідієносця або гіфи (рис. 15).
Недосконалі гриби поширені у природі. Більшість із них викликає різні захворювання? рослин і псування продуктів. Так, представники роду Рі~ запит є збудниками захворювання плодів та овочів (фузаріоз), викликають псування картоплі (суха гнилизна). Деякі види цього гриба виробляють отруйні в людини речовини, що викликають важкі харчові отруєння. Гриби роду ВоггуІз викликають псування цибулі, капусти, моркви, помідорів, а разом з іншими грибами - кагатну гниль цукрових буряків. Гриби роду А1-(егпапа вражають коренеплоди в період зберігання (чорна гчшль). Серцевину гниль буряків викликає гриб роду РЬо-та. Молочна цвіль ОеоШсЬіт сапсНйіт викликає псування квашених овочів, сметани, сиру та ін. із роду С1ас1озропіга часто виявляються на харчових продуктах, що зберігаються в холодильниках.
Дріжджі.Група дріжджів поєднує одноклітинні грибні організми, які мають справжнього міцелію.
Дріжджі широко поширені у природі. Вони мешкають в основному на рослинах, де є цукристі речі, які вони зброджують (нектар квітів, соковиті фрукти, ягоди, особливо перезрілі та пошкоджені, листя, стовбури берези під час скотини та дуба під час слизу, грунт). , дощ і комах.
Форма та розміри. Дріжджі можуть мати овальну, яйцеподібну, округлу, лимоноподібну, рідше - циліндричну, трикутну, серпоподібну, стрілоподібну, колбоподібну форми клітин. Розміри дріжджів варіюють у різних видів від 1,5 - 2 до 10 мкм у поперечнику і до 2-20 мкм (іноді до 50 мкм) в довжину.

Мал. 1.6. Схема будови дріжджової клітини:
1 - цитоплазматична мембрана; 2 - Клітинна стінка; 3 - Ядра; 4 - ядро; 5 - жирові краплі; 6 - мітохондрії; 7 – вакуоль; 8 - Гранули поліфосфату; 9 - ендоплазматична мережа; 10 - диктіосо-ми; 11 - нирковий рубець; 12 - рибосоми; 13 - цитоплазма
Деякі дріжджі на певній стадії розвитку можуть утворювати міцеліальні структури - псевдоміцелій.Дріжджі, як і гриби, є нерухомими організмами.
Будова клітини. Дріжджі, як і міцеліальні гриби, відносяться до еукаріотів і мають подібну до них будову-клітини, але є й деякі відмінності (рис. 16). Клітинна стінка дріжджів, на відміну грибів, на 60-70% складається з полісахаридів глюкану та маннану,пов'язаних з білками." і ліпідами, і лише невелика кількість (1-3%) складає хітин, який вкраплений у стінку у вигляді гранул. У ряду? дріжджів у певних умовах можуть утворюватися слизові капсули різної товщини полісахаридної природи. склеюватися один з одним, утворювати пластівці та осідати на дно судин, у яких вони розвиваються.
Клітини дріжджів, як і грибів, мають добре розвинений:; мембранний апарат – ЦПМ, ендоплазматичну мережу, апарат Гольджі, лізосоми, мітохондрії. У цитоплазмі є ядро. Рибосоми у дріжджів розташовуються в цитоплазмі та на? зовнішній стороні ядерної мембрани. Є вакуолі та включення запасних поживних речовин: ліпідів (особливо їх багато у дріжджів - продуцентів ліпідів), глікоген, метахро-матин. Клітинні структури дріжджів виконують ті ж функції, що і у грибів.
Розмноження та класифікація. Дріжджі розмножуються вегетативно та спорами, що утворюються безстатевим та статевим шляхом. Спосіб розмноження є важливим ознакою для класифікації дріжджів. До вегетативних способів розмноження відносяться: брунькування, поділ і поділ, що ниркується (рис. 17).
 |
Способи вегетативного розмноження дріжджів: брунькування; а- брунькування, б-розподіл; в -нирковий поділ
Ниркуваннянайбільш поширеним способом розмноження дріжджів. При брунькаванні на поверхні материнської (діляється) клітини виникає маленький горбок- нирка,яка поступово збільшується до до розмірів материнської клітини і перетворюється на дочірню клітину. Вона відокремлюється від материнської, залишаючи дома прикріплення нирковий рубець. На цьому місці нирка більше не утворюється. Може утворюватися одна нирка (полярна брунька), дві нирки на різних кінцях материнської клітини.(біполярна брунька), в декількох місцях поверхні материнської клітини (множина брунькування). Дочірні клітини можуть не відокремлюватися від материнської та залишатися з'єднаними з нею. Ниркування характерне для дріжджів овальної та округлої форми.
У деяких дріжджів при брунькаванні дочірні клітини не відокремлюються від материнської, а витягуються в довжину і продовжують утворювати все нові і нові нирки, що призводить до утворення хибного міцелію (псевдоміцелій). Псевдомі-аделій характерний для плівчастих дріжджів.
ПоділКлітини в результаті утворення в ній поперечної перегородки - септи - характерно для дріжджової циліндричної форми.
Ниркування розподілхарактеризується тим, що утворення дочірніх клітин починається з брунькування, а закінчується появою добре помітної септи в районі перешийка. Такий спосіб розмноження характерний для дріжджів лимонної форми.
Будь-якому вегетативному способу розмноження передує розподіл ядра, при якому одне з ядер, що знову утворилися, разом з цитоплазмою і частиною клітинних структур переходять вдочірню клітину і вони мають можливість самостійно існувати. У деяких дріжджів є спосіб безстатевого розмноження за допомогою безстатевих спор, що утворюються без злиття дріжджових клітин. Безстатеві суперечки - ендоспори - нерідко з'являються в невизначеному,числі в старих: культурах дріжджів, що розмножуються поділом і утворюють міцелій.
Статеве розмноження у дріжджів відбувається також за допомогою спор, але їх утворенню передує процес копуляції (злиття вмісту двох клітин та їх ядер). Утворюється зигота, у якій потім формуються суперечки: ядро ділиться, навколо нових ядер ущільнюється цитоплазма, і вони покриваються щільною оболонкою. Зигота зі спорами всередині 1 називається аском (сумкою), а суперечки-аскоспорами. Такі дріжджі відносяться до класу аскоміцетів, і їх називають аскоміцетовими дріжджами. Аскоспори можуть утворювати лише молоді клітини, вирощені на повноцінному живильному середовищі та перенесені до умов голодування, поганого постачання киснем та вологою. У різних видів дріжджів в асці утворюється 2-4, інколи ж 8 спор.
За сприятливих умов аскоспори виходять із аска і перетворюються на вегетативні клітини. У деяких видів дріжджів можуть зливатись ядра материнської та дочірньої клітин або ядра двох сестринських бруньок. Іноді відбувається копуляція сусідніх клітин, що проростають суперечка.
Аскоспори у дріжджів можуть бути овальними, круглими, бобоподібними, голкоподібними, шоломоподібними, кепкоподібними. з гладкою, зморшкуватою поверхнею, з бородавчастими або шилоподібними виростами і т. д. Спори дріжджів, так само як і суперечки міцеліальних грибів, виконують подвійну функцію: служать для перенесення несприятливих умов, але головне, на відміну від ендоспор бактерій, вони служать . Суперечки дріжджів більш стійкі, ніж вегетативні клітини, але менш стійкі, ніж бактеріальні суперечки. Так, спори дріжджів витримують нагрівання при температурі на 10 ° більше, ніж вегетативна клітина (40-50 ° С), а суперечки бактерій - на 50-60 ° С більше, ніж вегетативні клітини (60-120 ° С).
Оскільки дріжджі по суті є одноклітинними неміцеліальними грибами, вони включені до класифікації грибів. Однак в окрему систематичну одиницю вони не виділені, а розподілені за трьома класами грибів - аскоміцетів, базидіоміцетів та дейтероміцетів. Для мікробіології харчових виробництв мають значення лише аскоміцетові та недосконалі дріжджі. Між цими дріжджами є важлива різниця: у аскоміцетових дріжджів є. статевий процес і вони викликають енергійне спиртове бродіння. .Недосконалі дріжджі статевого процесу немає і, зазвичай, викликають слабке спиртове бродіння чи взагалі його викликають.
Аскоміцетові дріжджі. Включають приблизно 2/3 дріжджів. Серед них найбільше практичне значення мають цукроміцети, що поєднують більше половини відомих пологів-дріжджів. Особливо важлива роль належить роду цукроміцети, всі види якого викликають енергійне спиртове бродіння. Дріжджі цього роду розмножуються безстатевим способом (брунькування) і за допомогою аскоспор, які утворюють статевим шляхом.
У харчових виробництвах найбільш широко використовуються два види дріжджів цього роду: Сахароміцес церевізія (великі овальні клітини) у виробництві етилового спирту, пива, -квасу і в хлібопеченні і Сахароміцес еліпсойдес (великі еліптичні клітини) - їх використовують переважно у виноробстві. У кожному з цих виробництв застосовують свої, специфічні раси(різновиди) даних видів дріжджів, які мають найбільш цінні виробничі властивості.
До аскоміцетових дріжджів відносяться й інші пологи дріжджів. Це рід шизосахароміцети, клітини яких мають паличкоподібну форму і розмножуються розподілом або за допомогою «аскоспор, що утворюються в результаті статевого розмноження * (їх кількість 4-8). Дріжджі цього викликають спиртове бродіння. Вид З Шизосахароміцес помбе використовується вбродильної промисловості у країнах із спекотним кліматом, наприклад, в Африці, де виробляють пиво сорту «Помбе». Дріжджі роду Сахаромікоди мають великі клітини лимоноподібної форми. Вони розмножуються способом поділу, що нириться, на обох кінцях "клітини (біполярно) і за допомогою аскоспор (їх кількість 2-4), які розташовані парами і утворюються статевим шляхом. Причому при статевому способі відбувається копуляція спор в асці, а не злиття зкліток дріжджів. Ці дріжджі викликають спиртове бродіння, вони є шкідниками у виноробстві, тому що утворюють (продукти, що надають винам неприємний прокислий запах.
Деякі аскоміцетові дріжджі використовуються в мікробіологічній промисловості для одержання ліпідів та вітамінів. Так, дріжджі роду Ліпоміцес мають великі круглі клітини, які у старих культурах заповнені цілком великою краплею жиру. Зазвичай вони мають добре виражені капсули. Дріжджі роду Ліпоміцес розмножуються брунькуванням та аскоспорами, кількість яких у деяких видів може сягати 30 в одному асці.
Недосконалі дріжджі. Належать до класу дей-тероміцетів. Вони не утворюють суперечку, тому ці дріжджі часто називають аспорогенними.Розмножуються вони брунькуванням. Недосконалі дріжджі викликають або слабке бродіння, або не викликають його взагалі, тому часто називають нецукроміцетами.
Багато хто з них є причиною псування харчових продуктів і є шкідниками ряду харчових виробництв. Однак деякі з недосконалих дріжджів знайшли корисне практичне застосування. Серед недосконалих дріжджів найбільше значення мають пологи Кандіда, Торулопсіс та Родоторула.
Дріжджі роду Кандида мають подовжену форму клітин, поєднання яких утворюють примітивний псевдоміцелій. Багато хто з них не викликає спиртового бродіння і є шкідниками в бродильних виробництвах (наприклад, Кандіда мікодерму), оскільки, будучи аеробами, окислюють спирт до ді: оксиду вуглецю (вуглекислого газу) і води. Інші представники роду Кандида є шкідниками в дріжджовому виробництві, знижують якість хлібопекарських дріжджів, оскільки ставляться до видів, що слабо зброджують. Деякі з них викликають псування квашених овочів, безалкогольних напоїв та інших продуктів. Серед цих дріжджів є патогенні види, що викликають кандидози, що вражають слизові оболонки порожнини рота, носоглотки та інших органів людини. Різні види дріжджів роду Кандіда використовуються для одержання кормового білка та білково-вітамінних концентратів (БВК).
Дріжджі роду Торулопсіс мають дрібні круглі або овальні клітини. Багато видів здатні викликати слабке спиртове бродіння і використовуються у виробництві кефіру та кумису. Деякі застосовуються для промислового одержання кормового білка.
Дріжджі роду Родоторула мають круглі, овальні або подовжені клітини, останні утворюють псевдоміцелій. Колонії таких дріжджів червоні та жовті завдяки наявності пігментів каротиноїдів, які є провітаміном А. Ці дріжджі використовуються для промислового отримання кормових білково-каротиноїдних концентратів, які є джерелом жиророзчинного вітаміну А для тварин. Інші представники цього роду накопичують у клітинах багато ліпідів і використовують у мікробіологічної промисловості як продуценти ліпідів поруч із представниками недосконалих дріжджів іншого роду - Криптококкус.
ВІРУСИ
Винахід електронного мікроскопа дозволило вперше спостерігати найдрібніші організми - віруси та фаги. Віруси часто називають такими, що фільтруються за їх здатність проходити через пори бактеріологічних фільтрів, які затримують бактерії при механічному способі стерилізації. Віруси було відкрито 1892 р. російським ботаніком Д. І. Івановським щодо хвороби тютюну - тютюнової мозаїки. Їхні розміри коливаються від 10-12 нм (віруси ящуру, поліомієліту) до 200-350 нм (віруси віспи, герпесу).
Віруси немає клітинного будови. Вони бувають кулястої, паличкоподібної, ниткоподібної та сперматозоїдної форми. Вірусна частка називається віріоном. Вона складається з нуклеїнової кислоти (ДНК або РНК) та білка глобуліну; деякі віруси містять також ліпіди та вуглеводи. Харак-

Мал. 18. Схема будови фага:
1 - Головка; 2 - ДНК; 3 - відросток; 4 - стрижень; 5 - базальна пластинка з шипами; 6 - нитки відростка
терною особливістю вірусів є їх здатність до утворення кристалів, що тривалий час служило причиною суперечок про живу або неживу природу вірусів. Згодом було доведено, що кристали – це нуклеїнова кислота та білок. Потім було встановлено ряд властивостей, що підтвердили уявлення про живу природу вірусів, - здатність до самовідтворення (розмноження), мінливість, пристосовність до умов існування, а також здатність викликати інфекційні процеси. Розвиток і розмноження вірусів можливе лише у клітинах живого організму - господаря, тобто є паразитами людини, викликаючи інфекційні захворювання (грип, поліомієліт, кір, вітряна віспа та інших.), і навіть тварин і рослин.
Для лікування деяких захворювань, викликаних вірусами грипу, герпесу та аденовірусами, застосовують ферментні препарати- нуклеази,викликають руйнування нуклеїнових кислот, що позбавляє віруси здатність до самовідтворення, а отже, ліквідує їх інфекційність.
У 1898 р. російський учений Н. Ф. Гамалея щодо сибірки великої рогатої худоби вперше спостерігав, що спорообразующие палички - збудники хвороби - розчиняються під впливом якогось агента. У 1915 р. англійським мікробіологом Ф. Туортом і в 1917 р. канадським мікробіологом Ф. Д "Еррелем була встановлена природа цього явища. Воно отримало назву бактеріофагії, а збудник - бактеріофага («пожирач бактерій»).
Розміри фагів коливаються від 40 до 140 нм. Бактеріофаги мають вигляд багатогранної головки зі стрижнем,покритою зовні білковою оболонкою (рис. 18). Усередині стрижня є канал. Головка фага заповнена молекулою ДНК. В основі стрижня є базальна платівка зшипами та нитками.
Вплив фага на бактеріальну клітину відбувається в кілька стадій (рис. 19): адсорбція фага на бактеріальній клітині за допомогою базальної платівки із зубцями та нитками, проникнення ДНК з головки фага по каналу в бактеріальну клітину, в якій потім під впливом фагової ДНК

Рис* 19. Схема розвитку фага в бактеріальній клітині:
а -адсорбція; б- перехід ДНК у клітину; в- перебудова обміну речовин у клітині;
г -освіту нових частинок бактеріофага; д -розчинення клітинної стінки
відбувається повна перебудова обміну речовин, синтезується вже не бактеріальна ДНК, а фагова, що наводить доутворенню в бактеріальній клітині нових частинок фага розчинення клітинної стінки бактерії, її загибель.
Бактеріофаги завдають великої шкоди у молочній промисловості (виробництві сирів, сиру, сметани) та у виробництві маргарину. Вони вражають в основному молочнокислі стрептококи заквасок для отримання цих продуктів. Під впливом бактеріофага клітини стрептококів лізуються (розчиняються) та гинуть. В антибіотичній промисловості актинофаги лізують виробничу культуру актиноміцетів – продуцентів антибіотиків.
У медицині бактеріофаги використовуються для лікування деяких захворювань, наприклад дизентерії.
Вступ
Мікробіологія - наука про дрібні, невидимі неозброєним оком організми, названі мікробами або мікроорганізмами. Вона вивчає закономірності їх життя та розвитку, а також зміни, які вони викликають в організмі людей, тварин, рослин і в неживій природі. Розвиток мікробіології, як та інших наукових дисциплін, перебуває у тісній залежності від способів виробництва, запитів практики, загального прогресу науку й техніки.
Метою мікробіології як науки є вивчення систематики, морфології (форми та будови) та фізіології (життєдіяльності) мікроорганізмів, методів їх виділення та розпізнавання, а також з'ясування їх значення у природі та можливостей застосування у різних сферах діяльності людини.
Мікробіологічний контроль на харчових виробництвах є всі методи дослідження та контролю, пов'язані з визначенням ступеня бактеріальної обсімененості контрольованого об'єкта, а також методи кількісного обліку мікрофлори.
Морфологія бактерій
Форма бактерій
Переважна більшість відомих бактерій мають форму або сфери (кулясті), або циліндра (паличкоподібні), або спіралі. Кулясті бактерії (рис. 1.) бувають одиночними (коки), з'єднаними по дві клітини (диплококи), по чотири клітини (тетракоки), у довгі ланцюжки (стрептококи), у пакети (сарцини), у вигляді скупчень неправильної форми (стафілококи) . Паличкоподібні бактерії (рис. 1) відрізняються за величиною відношення довжини клітини до її поперечного розміру. У коротких паличок це ставлення так мало, що їх важко відрізнити від коків вони поділяються на бактерії (що не утворюють суперечки) та бацили (утворюючі суперечки). Бактерії спіралеподібної форми характеризуються різним числом витків спірили мають від одного до декількох витків, вібріони виглядають як вигнуті палички, їх можна розглядати як неповний виток спіралі.
З розвитком мікроскопічної техніки та удосконаленням методів підготовки препаратів відкрито інші екзотичні форми бактерій. Деякі бактерії мають вигляд зімкнутого або розімкнутого кільця, у деяких видно клітинні вирости (простеки), кількість яких коливається від 1 до 8 і більше, виявлено бактерії червоподібної форми, схожі на кристали, і т.п.
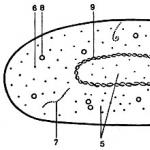
Будова бактеріальної клітини
Бактеріальна клітина має дуже складну гетерогенну і водночас суворо впорядковану структуру. Загалом будова бактеріальної клітини не відрізняється від будови клітини вищих організмів. Клітина як універсальна одиниця життя виявилася настільки досконалою формою організації живої матерії, що у процесі еволюції від одноклітинних до вищих багатоклітинних організмів вона зберегла основні риси своєї будівлі, отже, і функції.
Малюнок 1. Форми бактерій. Кулясті: а - мікрококи, б - стрептококи, в - диплококи, г - стафілококи, д - сарцини; паличкові: е - бактерії, ж - бицили, з, і - звивисті, до - спірили.
На рис. 2. представлена схема будови бактеріальної клітини відомого цитолога В.І. Бірюзовий. Форма бактеріальної клітини визначається жорсткою (ригідною) клітинною стінкою, яка надає клітині певної, спадково закріпленої зовнішньої форми. На клітинній стінці бактерій перебувають звані поверхневі структури: капсула, джгутики, статеві ворсинки, вії. Під клітинною стінкою розташована цитоплазматична мембрана (ЦПМ), яка відмежовує цитоплазму клітини. Цитоплазматична мембрана разом із цитоплазмою називається протопластом. Усі шари, що розташовуються із зовнішнього боку від цитоплазматичної мембрани, називають клітинною оболонкою.
Клітинна стінка.
У прокаріотів клітинна стінка складається з пептидоглікану, якого немає в еукаріотних клітинах. Залежно від будови клітинної стінки прокаріоти поділяються на дві групи: грампозитивні та грамоздатні. Такий підрозділ заснований на відмінності у способі забарвлення, запропонованому в 1884 датським ученим X. Грамом. Клітинні стінки грампозитивних і грамнегативних прокаріотів різко різняться як по хімічному складу, так і по ультраструктурі.

Мал. 2. Схема будови бактеріальної клітини: Про - оболонка клітини; ЦМ – цитоплазматична мембрана; М – мітохондрія (мезосома); Ж-жирові включення; ЯВ – ядерна вакуоль; ДНК – нитки ДНК; ЕС – ергастоплазматична система; Р – рибосоми; В – волютин; Г - глікоген
У клітинній стінці грампозитивних бактерій міститься 50...90% пептидоглікану, грамнегативних бактерій - 1...10% пептидоглікану. Крім пептидоглікану в клітинній стінці грампозитивних прокаріотів містяться унікальні хімічні сполуки - тейхоєві кислоти. На клітинну стінку прокаріот припадає від 5 до 50 % сухих речовин клітини.
Клітинна стінка прокаріотів виконує різноманітні функції: механічно захищає клітину від впливів навколишнього середовища, забезпечує підтримку її зовнішньої форми, дає можливість клітині існувати в гіпотонічних розчинах. У клітинній стінці розташовані канали, або дифузійні пори, для пасивного транспортування речовин та іонів у клітину.
Клітинна стінка перешкоджає проникненню в клітину токсичних речовин. На зовнішній стороні клітинної стінки розташовано багато макромолекул, що контактують із навколишнім середовищем: специфічні рецептори для фагів, антигени, макромолекули, що забезпечують міжклітинні взаємодії при кон'югації, а також між патогенними бактеріями та клітинами та тканинами вищих організмів.
Поверхневі структури.
У бактерій зовні клітинної стінки є капсула (рис. 3.) - Слизова освіта, що обволікає клітину, що зберігає зв'язок з клітинною стінкою і має аморфну будову. Залежно від товщини капсули бувають мікрокапсули (товщина менше 0,2 мкм) та макрокапсули (товщина більше 0,2 мкм). Капсули захищають клітину від механічних пошкоджень, висихання, створюють додатковий осмотичний бар'єр, є перешкодою для проникнення фагів. Іноді капсула є джерелом запасних поживних речовин. Слиз допомагає прикріпленню клітин до різних поверхонь. В даний час здатність деяких бактерій синтезувати капсули (своєрідні позаклітинні полімери) використовують на практиці як замінники плазми крові та для отримання синтетичних плівок.
Рис.
Багато бактерій нерухомі, якщо вони здатні пересуватися, це рух забезпечується джгутиками - структурами, розташованими лежить на поверхні клітин. Число, розміри та розташування джгутиків, як правило, є ознакою, постійною для даного виду (рис. 4.), і мають таксономічне значення. Без джгутиків здатні пересуватися лише ковзні бактерії та спірохети. Зазвичай товщина джгутика становить 15-20 нм, довжина 3-15 мкм. Бактерії зі джгутиками можуть рухатися дуже швидко, наприклад, Bac. megaterium зі швидкістю 16 мм/хв, Vibro cholerae – 12 мм/хв.

Мал. 4.
При полярному розташуванні джгутиків вони діють подібно до корабельного гвинта і проштовхують клітину в навколишньому рідкому середовищі. Обертальний рух джгутика відбувається за рахунок базального тіла. Джгутики обертаються порівняно швидко. У спірил вони роблять близько 3000 об/хв, що близько швидкості середнього електромотора. Обертання джгутиків викликає і обертання клітини з 1/3 цієї швидкості у протилежному напрямку.
Перитрихіально розташовані джгутики Е. coli працюють як один добре скоординований спіральний пучок і проштовхують клітину через довкілля (рис. 4.).
Вивчення джгутиків в електронному мікроскопі показало, що вони складаються із трьох частин (рис. 5.). Основну масу джгутика становить довга спіральна нитка (фібрила), що переходить біля поверхні клітинної стінки в потовщену вигнуту структуру - гачок. Нитка за допомогою гачка прикріплена до базального тіла, яке є системою з двох або чотирьох кілець (L, Р, Sw. M), нанизаних на стрижень, що є продовженням гачка.

Мал. 5.
Останнім часом досягнуто великих успіхів у розшифровці механізму руху прокаріотів. Прокаріотна клітина має механізм, що дозволяє перетворювати електрохімічну енергію безпосередньо на механічну. Крім джгутиків на клітинній стінці прокаріотної клітини можуть бути статеві ворсинки та вії у вигляді різної довжини виростів (рис. 6).

Мал. 6. Типи волосин E.coli: F - джгутики, S - статеві ворсинки (F-like sexpili), C - вії.
Цитоплазматична мембрана.
Під клітинною стінкою розташована цитоплазматична мембрана, яка є обов'язковим структурним елементом будь-якої клітини, порушення цілісності якого призводить до втрати клітиною життєздатності. Перед ЦПМ припадає 8... 15 % сухої речовини клітини. ЦПМ - це білково-ліпідний комплекс та невелика кількість вуглеводів.
ЦПМ виконує різноманітні функції за допомогою спеціальних переносників, які називають транслоказами. Через мембрану здійснюється спеціальне перенесення різних органічних та неорганічних молекул та іонів.
У ЦПМ локалізовано багато ферментів. ЦПМ є основним бар'єром, що забезпечує вибіркове надходження в клітину та вихід з неї різноманітних речовин та іонів.
У прокаріотів описані локальні вп'ячування ЦПМ, які називаються мезосомами. Мезосоми розрізняються розмірами, формою та локалізацією у клітині. Вважається, що з мезосом пов'язане посилення енергетичного метаболізму клітин.
цитоплазма.
Вміст клітини, оточений ЦПМ, називається цитоплазмою. Цитоплазма має гомогенну консистенцію та містить набір розчинних РНК, ферментів, продуктів та субстратів метаболічних реакцій. У цитоплазмі розташовані різноманітні структури: рибосоми, генетичний апарат (ДНК) та включення різної хімічної природи та функціонального призначення.
Рибосоми - рибонуклеопротеїдні частки розміром 15-20 нм. Їхнє число в клітині залежить від інтенсивності процесу синтезу білка. У клітині, що швидко росте, Escherichia coli міститься приблизно 15000 рибосом. Синтез білка здійснюється агрегатами, що складаються з рибосом, інформаційних молекул і транспортних РНК, званих полірибосомами. Генетичний апарат прокаріотної клітини представлений однією молекулою ДНК, що має форму ковалентно замкнутого кільця та назва бактеріальної хромосоми. Довжина молекули ДНК у розгорнутому вигляді може становити понад 1 мм, тобто. майже 1000 разів перевищувати довжину бактеріальної клітини. Генетичний апарат прокаріотної клітини називають нуклеоїдом.
У цитоплазмі прокаріотів розташовані різні включення, частина яких виконує функцію запасних поживних речовин, представлених полісахаридами, ліпідами, поліпептидами, поліфосфатами, відкладеннями сірки. Полісахариди – це глікоген, крохмаль, гранульоз (крохмалоподібна речовина). Найчастіше зустрічаються у представників анаеробних спорових бактерій групи клостридіїв. У несприятливих умовах вони використовуються як джерела вуглецю та енергії. Ліпіди накопичуються у вигляді гранул, що складаються з полімеру -оксимасляної кислоти. У деяких прокаріотів, що окислюють вуглеводні, полі-в-оксимасляна кислота становить до 70% сухої речовини клітини.
Ліпіди служать для клітини добрим джерелом вуглецю та енергії. Поліфосфати, що також накопичуються у вигляді гранул, називаються волютиновими та використовуються клітинами як джерело фосфору. Для бактерій, що здійснюють хемосинтез за рахунок окислення сірководню, характерне накопичення у клітинах молекулярної сірки. Усі накопичені (запасні) речовини, які представлені у вигляді високомолекулярних полімерних молекул, відмежовані від цитоплазми білковою мембраною.
Пігменти бактерій
Колонії багатьох бактерій яскраво забарвлені. Здатність синтезу пігментів зумовлена генетично. Серед пігментів бактерій зустрічаються каротиноїди, феназинові барвники, піроли, азахінони, антоціани та ін.
Пігменти захищають клітини від світлового ушкодження та використовують світло для фотосинтезу. У багатьох мікроорганізмів утворення пігменту йде лише на світлі. Наприклад, яскраво-червоне забарвлення колоній у Serratia marcescens обумовлено присутністю пігменту продігіозину. Бактерії Pseudomonas indigofem, Cotynebacterium insidiosum, Arthrobacter atrocyaneus та ін синтезують індигоїдин - нерозчинний у воді синій пігмент, що виділяється в середу. Chromobacterium violaceum утворює синьо-фіолетовий пігмент віолацеїн, нерозчинний у воді. Віолацеїн є похідним індолу, що утворюється при окисненні триптофану. Pseudomonas aeruginosa утворює пігмент нікс-піоціаніну. Різні штами псевдомонаду утворюють такі пігменти, як Феназин-1-карбонову кислоту, оксихлорофін, йодинін, а іноді всі пігменти разом.
Усі пігменти ставляться до вторинних метаболітів, тобто. вони не належать до тих сполук, які є у всіх організмів і є похідними звичайних метаболітів або структурних компонентів клітини. Деякі пігменти мають антибіотичні властивості, так що багато пігментованих мікроорганізмів є продуцентами антибіотиків.
Зростання та способи розмноження бактерій
Зростання прокаріотної клітини – це узгоджене збільшення кількості всіх хімічних компонентів, з яких вона побудована. Зростання є результатом безлічі скоординованих біосинтетичних процесів, що знаходяться під строгим регулярним контролем, і призводить до збільшення маси та розмірів клітини. Зростання клітини не безмежне. Після досягнення певних (критичних) розмірів клітина піддається поділу. Для більшості прокаріотів характерний рівновеликий бінарний поперечний поділ, що призводить до утворення двох однакових дочірніх клітин.
У більшості грампозитивних бактерій розподіл відбувається шляхом синтезу поперечної перегородки, що йде від периферії до центру. Поперечна перегородка формується з ЦПМ та пептидогліканового шару. Зовнішні шари синтезуються пізніше.
Клітини більшості грамнегативних бактерій діляться шляхом перетяжки. Наприклад, у Е. coli на місці розподілу виявляється поступово збільшується і спрямоване всередину викривлення ЦПМ та клітинної стінки.
Варіантом бінарного поділу є брунькування, у якому одному з полюсів материнської клітини утворюється невеликий виріст (нирка), що збільшується у процесі зростання. Поступово нирка досягає розмірів материнської клітини та відокремлюється від неї. Клітини, що брунькуються, піддаються старінню. При рівновеликому бінарному поділі материнська клітина дає початок двом дочірнім клітинам, а сама зникає. При брунькаванні материнська клітина дає початок дочірній клітині і з-поміж них можна знайти морфологічні відмінності. Розподіл прокаріотної клітини починається, як правило, через деякий час після завершення циклу поділу ДНК.
Спороутворення бактерій
Вегетативні клітини багатьох прокаріотів для перенесення несприятливих умов утворюють спеціальні клітини (ендоспори), що мають підвищену стійкість. p align="justify"> В основі морфологічного диференціювання лежать біохімічні процеси, запрограмовані відповідною генетичною інформацією. Освіта ендоспор відбувається у прокаріотів та грибів.
Ендоспора формується всередині материнської клітини (спорангія), має специфічні структури: багатошарові білкові покриви, зовнішню і внутрішню мембрани, кортекс (рис. 7.). Ендоспори стійкі до підвищених температур, доз радіації, які детальні для вегетативних клітин. До спороутворюючих бактерій належить велика кількість прокаріотів з 15 пологів, серед яких є паличкоподібні, сферичні, спірили та нитчасті організми. Всі вони мають клітинну стінку, характерну для грампозитивних прокаріотів. У кожній бактеріальній клітині утворюється, як правило, одна ендоспора.
Найкраще процес спороутворення вивчений у представників пологів Bacillus та Clostridium. Перед спороутворенням відбувається поділ ДНК вегетативної клітини. Утворюється тяж уздовж довгої осі клітини, потім приблизно 1/3 тяжа відокремлюється і переходить у суперечку, що формується, в одному з полюсів клітини. Потім відбувається ущільнення цитоплазми, яка разом із ДНК відокремлюється від решти вмісту клітин за допомогою перегородки. Перегородка формується шляхом вп'ячування ЦПМ від периферії до центру, де зростається, і утворюється спорова оболонка. Відсічена ділянка «обростає» другою мембраною і утворюється проспора. На наступному етапі між мембранами проспори починає формуватися кортекс, а зовні синтезуються спорові покриви, які з кількох шарів.

Мал. 4 а і б -освіта септи, в і г -оточення протопласту суперечки протопластом материнської клітини, д утворення кортексу і оболонок спори; е-схема будови зрілої суперечки: 1 - екзоспоріум, 2 - зовнішня оболонка суперечки; 3 – внутрішня оболонка суперечки; 4 – кортекс; 5-клітинна стінка зародка, 6-цитоплазматична мембрана, 7 - цитоплазма з ядерною речовиною
У багатьох бактерій поверх покривів ендоспори формується ще одна структура - екзоспоріум, структура якого залежить від виду бактерій. У прокаріотів з роду Clostridium виявлені придатки на екзоспоріумі різної будови, іноді дуже химерної (рис. 8). Функціональне значення цих виростів не з'ясовано.
Ендоспори прокаріотів характеризуються дуже низьким рівнем метаболізму, вони дуже стійкі до впливу факторів зовнішнього середовища: високих і низьких температур, зневоднення, літичних факторів, високої кислотності середовища, радіації, механічних впливів тощо. Механізми стійкості ендоспор поки що мало вивчені. Вважається, що ендоспорам прокаріотів надають стійкості зневодненого стану цитоплазми, термостійкості спорових ферментів, наявності дипіколінової кислоти та великої кількості двовалентних катіонів. Стійкості ендоспор сприяють також поверхневі структури: мембрани, кортекс, покриви, що механічно захищають вміст ендоспор від проникнення ззовні агресивних речовин.

Сформовані ендоспори, що покоїться, можуть перебувати в життєздатному стані протягом різного часу: від декількох діб до 1000 років і більше. Нижче наводиться залежність виживання спор різних груп бактерій від ушкоджуючих факторів (високої температури та висушування).
Таблиця 1

При мікроскопічному дослідженні ендоспори добре помітні. У сумнівних випадках можна використовувати спеціальне фарбування, для чого фіксований препарат треба прокип'ятити з карболовим розчином фуксину. Ендоспори міцно пов'язують барвник і не знебарвлюються навіть при обробці етанолом або оцтовою кислотою; весь інший вміст клітини при цьому знебарвлюється.
Ендоспора містить майже всю суху речовину клітини, але займає в 10 разів менший обсяг. Ендоспори є обов'язковою стадією життєвого циклу бацил. За сприятливих умов харчування бацили можуть необмежений час розмножуватися поділом. Утворення ендоспор починається тільки тоді, коли не вистачає поживних речовин і коли надміру накопичуються продукти обміну.
За сприятливих умов більшість ендоспор проростає. Відсоток проростання ендоспор можна збільшити, прогрів одні суперечки у воді при 60 ° С протягом 5 хв, інші - при 100 ° С протягом 10 хв. Тепловий шок повинен проводитись безпосередньо перед висівом.
У харчовій промисловості для знищення термостійких ендоспор бактерій вдаються до дорогої стерилізації харчових продуктів. Наприклад, якщо при пастеризації (нагрівання при 80 ° С протягом 10 хв) харчових продуктів вегетативні клітини спороутворюючих бактерій та решта бактерій гинуть, то термостійкі ендоспори витримують значно сильніше нагрівання, а деякі суперечки - навіть кип'ятіння протягом декількох годин.
Класифікація бактерій
У міру опису все нових бактерій з'явилася гостра необхідність систематизувати і порівнювати з відомими культури, що знову описуються. Наприкінці XIX – на початку XX ст. з'явилися визначники бактерій, які допомагають класифікувати, ідентифікувати знову виділені бактерії за певними ознаками. p align="justify"> При класифікації основним завданням є визначення виду бактерій.
Вид - це група близьких між собою організмів, що мають однакове походження і характеризуються певним і морфологічними, біохімічними та фізіологічними ознаками, що сприяють пристосуванню до певного довкілля.
Види об'єднуються у пологи, пологи - у сімейства, потім слідують порядки, класи, відділи, царства. Вид бактерій описують з допомогою ознак: морфологічних, культуральних, фізіолого-біохімічних та інших.
Морфологічними ознаками є форма клітини, наявність або відсутність джгутиків, капсули, здатність до спороутворення, забарвлення за Грамом.
До культуральних ознак належать загальний вигляд бактеріальної колонії, наявність пігменту та ін.
Фізіолого-біохімічними ознаками є спосіб отримання енергії, потреби в поживних речовинах, ставлення до факторів довкілля та ін.
Найсучаснішим визначником для ідентифікації бактерій є «Короткий визначник бактерій Берги», що найбільш повно описує відомі бактерії. У 8-му виданні цього визначника всі бактерії, крім ціанобактерій, згруповані в 19 частинах. Нижче наводиться їх коротка характеристика:
Частина 1. Фототрофні бактерії. У цій частині згруповані фотосинтезуючі бактерії, що характеризуються специфічним набором пігментів та особливим типом фотосинтезу: пурпурові бактерії та зелені серобактерії. Пігменти представлені різними видами бактеріохлорофілу та каротиноїдами. Фотосинтез не супроводжується виділенням кисню. Це переважно водні мікроорганізми.
Частина 2. Ковзаючі бактерії. До складу бактерій цієї частини віднесено два порядки: міксобактерії (Mixobacteriales) та цитофаги (Cytoppagales). До першого порядку відносяться бактерії, що утворюють шар слизу навколо клітки. Бактерії рухливі. Міксобактерії утворюють так звані плодові тіла, всередині яких клітини переходять у стан, що покоїться. До другого порядку відносяться бактерії, за типом руху подібні до міксобактерій, але не утворюють плодових тіл. До складу порядку входить чотири сімейства переважно водних бактерій.
Частина 3. Хламідобактерії. До складу цієї частини входять ниткоподібні бактерії, оточені загальною піхвою, слизовою оболонкою. Піхва складається з гетерополісахариду, часто інкрустованого окислами заліза або марганцю. Зустрічаються у водоймах та ґрунті.
Частина 4. Ниркові та (або) стеблинні бактерії. До складу цієї частини входять бактерії, що утворюють придатки (стеблинки), що складаються зі слизу і не пов'язані з цитоплазмою клітини, а також бактерії, що утворюють ниткоподібні клітинні вирости - простіки. Бактерії широко поширені у ґрунті та водоймах.
Частина 5. Спірохети. Ця частина поєднує бактерії, що мають вигляд тонких спіралеподібних одноклітинних форм. Багато бактерій патогенні, викликають сифіліс, зворотний тиф.
Частина 7. Грамнегативні аеробні палички та коки. У цю частину бактерій входять п'ять сімейств, одне з яких - Pseudomonas - поширене в природі: у повітрі, грунті, морських і прісних водах, мулах, стічних водах, в харчових продуктах. В останніх бактерії цього сімейства викликають псування.
Частина 8. Грамнегативні факультативно-анаеробні палички. До складу цієї частини входять два сімейства: ентеробактеріацеа (Enterobacteriaceae) та вібріонацеа (Vibrionaceae). Ентеробактеріацеа є Грамнегативними, безспоровими, аеробними або факультативно-анаеробними паличками. Найбільш вивченими представниками цього сімейства є бактерії Escherichia coli, які завжди містяться в кишечнику людини та тварин, тому про забруднення води та харчових продуктів судять за наявністю в них Е. coli. E. coli належать до умовнопатогенних бактерій. До збудників тяжких кишкових захворювань людини належать бактерії цього сімейства з пологів сальмонела (Salmonella) та шигела (Shigella). Бактерії Salmonella typhi є збудниками черевного тифу. Бактерії роду Shigella – збудники бактеріальної дизентерії. До сімейства вібріонацеа (Vibrionaceae) належать бактерії виду Vibrio cholerae – збудники азіатської холери.
Частина 9. Грамнегативні анаеробні бактерії. Бактерії, що згруповані в цій частині, відносяться до сімейства Bacteroidaceae. Всі вони є паличками; це облігатні анаероби. Основне місце проживання цих бактерій - кишечник людини і тварин, травний тракт комах. Деякі види є патогенними та викликають різні ураження шкіри, низки органів та тканин тіла.
Частина 13. Метанутворюючі бактерії. Ця частина представлена одним сімейством - Methanobacteriaceae. Усі бактерії однорідні за фізіологічними ознаками: це облігатні анаероби; Основний продукт енергетичного обміну - метан. Основні місця проживання - болота, різні очисні споруди, рубець жуйних тварин.
Частина 14. Грампозитивні коки. До цієї частини належать дві групи. Перша група - аеробні та (або) факультативноанаеробні бактерії сімейств Micrococcaceae та Streptococcaceae. Друга група - облігатні анаероби сімейств Peptococcaceae. Бактерії сімейства Micrococcaceae - це коки, що діляться більш ніж в одній площині, іноді не розходяться, утворюючи скупчення сферичної або неправильної форми. Енергію одержують за рахунок дихання або бродіння. В основному це сапрофіти, що руйнують багато складних органічних речовин і виконують функцію «сміттярів». Багато з них є збудниками псування харчових продуктів. Серед них є патогенні форми, що належать до роду Staphylococcus. Розвиваючись на харчових продуктах, виробляють токсини, які викликають отруєння.
Бактерії сімейства Streptococcaceae - це коки, нерухомі, безспорові, факультативно-анаеробні. Бактерії пологів Streptococcus, Pediococcus, Aerococcus є гомоферментативними молочнокислими; Бактерії роду Leuconostoc - гетероферментативні молочнокислі.
Бактерії сімейства Peptococcaceae - це облігатно-анаеробні коки, що мешкають у ґрунті, на поверхні злаків, у ротовій порожнині, шлунково-кишковому тракті, дихальних шляхах людини та тварин; деякі види є патогенними.
Частина 15. Палички та коки, що утворюють ендоспори. Ця частина представлена одним сімейством - Bacillaceae, до складу якого входить п'ять пологів. Два з них - бацили (Bacillus) і клостридії (Clostridium) - найбільш численні і становлять найбільший інтерес. Бацили - це паличкоподібні бактерії; більшість із них рухливі; утворюють ендоспори; облігатні чи факультативні аероби. Бацили синтезують різні літичні ферменти, що розщеплюють білки, жири, полісахариди та інші макромолекули. Деякі види утворюють антибіотики. Більшість сапрофітів. Основним місцем проживання є ґрунт. Багато бацили є збудниками псування харчових продуктів. Серед них є патогенні для людини та тварин види, наприклад Bacillus anthracis – збудник сибірки.
До складу роду Clostridium входять палички, що відрізняються від бацил формою спороутворення та облігатно-анаеробним способом існування. Викликають маслянокисле бродіння, більшість клостридій - сапрофіти, жителі ґрунту. Деякі види живуть у кишечнику людини та тварин, наприклад Cl. teteni - збудник правця, Cl. perfringens - збудник газової гангрени, CL botulinum - продуцент екзотоксин, одного з найсильніших біологічних отрут.
Частина 16. Грамопозитивні аспорогенні паличкоподібні бактерії. Ця частина також представлена одним сімейством - Lactobacillaceae, до складу якого входить один рід - Lactobacillus. Бактерії отримують енергію за рахунок гомоферментативного або гетероферментативного молочнокислого бродіння, широко поширені в природі: в грунті, на залишках тваринного і рослинного походження, що розкладаються, в кишечнику хребетних, в молоці, молочних продуктах. Зустрічаються патогенні форми. Багато бактерій використовуються при виготовленні кисломолочних продуктів, сирів, квашення овочів, тіста та ін.
Частина 17. Актиноміцети та споріднені організми. Ця частина поєднує коринебактерії, пропіоново-кислі бактерії та актипоміцети.
З існуючих видів пропіоново-кислих бактерій найбільший інтерес у мікробіології продовольчих товарів представляє вид Propionibacterium freudenreichii роду Propionibacterium. Бактерії цього роду - грампозитивні нерухомі палички, що не утворюють суперечки. Це облігатні анаероби. Пропіоновокислі бактерії широко використовуються при виробництві сирів.
До актиноміцетів відносяться бактерії, що утворюють розгалужені нитки, іноді розвинений міцелій. Мають різні методи розмноження. Більшість актиноміцетів розмножуються за допомогою спор, що утворюються на спорангіях, які можуть бути довгі або короткі, прямі або спіралеподібні з різним числом завитків і розташуванням. Актиноміцети представлені двома сімействами: Mycobacteriaceae та Streptomycetaccae. Сімейство Mycobacteriaceae представлене одним родом - Mycobacterium, характерною ознакою якого є утворення розгалужених форм у молодому віці.
Більшість мікобактерій є сапрофітами, живуть у ґрунті та використовують найрізноманітніші органічні сполуки: білки, вуглеводи, жири, воску, парафіни. Деякі види патогенні, наприклад М. tuberculosis – збудник туберкульозу, М. leprae – збудник прокази.
Представники сімейства Streptomycetaceae утворюють добре розвинений повітряний міцелій, розмножуються спорами, що формуються на кінцях гіф, шматочками міцелію. Їх близько 500 видів. Багато стрептоміцети синтезують антибіотики, які проявляють активність проти бактерій, грибів, водоростей, найпростіших, фагів і мають протипухлинну дію.
Частина 18. Ріккетсії. До складу цієї частини входять два порядки бактерій - рикетсії (Rickettsiales) та хламідії (Chlamydiales).
Ріккетсії - це нерухомі бактерії, грамнегативні розмножуються тільки в клітинах господарів, викликаючи рикетсіози. Існують непатогенні види.
Частина 19. Мікоплазми. До мікоплазм відносяться прокаріоти у яких відсутня клітинна стінка, вони обмежені одне тришаровою мембраною. Клітини дуже дрібні, іноді ультрамікроскопічні, плеоморфні. Спосіб розмноження недостатньо зрозумілий; мабуть, відбувається за рахунок утворення кокоподібних структур «елементарних тілець», можливе біпарне поділ та розмноження брунькуванням. Стадії спокою невідомі. За обсягом генетичної інформації, що міститься в геномі, мікоплазми займають проміжне положення між Е. coli та Т-фагами.
Мікробіологія- це розділ біології, що вивчає закономірності життя та розвитку мікроорганізмів у єдності з навколишнім середовищем.
Ця наука вивчає властивості мікроорганізмів та процеси, які вони викликають у макроорганізмі.
Мікробіологія поділяється на розділи: Загальну та медичну. Медична на загальну, приватну та санітарну.
Загальна мікробіологіявивчає будову та життєдіяльність мікроорганізмів, спадковість, мінливість, фізіологію, цикли розвитку.
Медична мікробіологіявивчає патогенні мікроорганізми, що викликають захворювання у людини.
Мікроорганізми, що не викликають захворювання, називаються – сапрофіти.Існують захворювання, викликані умовно-патогенною мікрофлорою, захворювання розвиваються при попаданні мікроорганізмів, у певну не властиву для них, довкілля.
Приватна мікробіологіявивчає конкретних збудників захворювань та методи діагностики.
Медична мікробіологія пов'язана з іншими медичними дисциплінами, такими як інфекційні хвороби, епідеміологія, загальна гігієна, генетика, анатомія та фізіологія людини, основи сестринської справи, латинська та інші.
Чотири царства життя.
Світ мікроорганізмів різноманітний. У міру їх відкриття та вивчення мікроорганізми були розподілені на групи:
1. Бактерії
2. Променисті гриби
3. Нитчасті гриби
4. Дріжджові гриби
5. Синьо-зелені водорості
6. Спірохети
7. Найпростіші
8. Ріккетсії
9. Мікоплазми
10. Віруси
11. Плазміди
Єдине, що їх поєднує, - мікроскопічні розміри. Ці організми відрізняються один від одного за багатьма ознаками: за рівнем організації геномів, наявністю та складом білоксинтезуючих систем і клітинної стінки.
Відповідно до цих ознак всі живі істоти діляться на 4 царства: еукаріоти, прокаріоти, віруси та плазміди. До прокаріотів відносять бактерії, синьо-зелені водорості, спірохети, актиноміцети, рикетсії, мікоплазми. Найпростіші, нитчасті гриби та дріжджі-еукаріоти.
Відмінні риси перерахованих царств життя такі:
Прокаріоти -це організми, у яких немає оформленого ядра, а є лише попередник – нуклеоїд. Він представлений 1 або декількома хромосомами, які складаються з ДНК і вільно розташовуються в цитоплазмі, не відмежовані від неї ніякою мембраною. Прокаріоти не мають апарату мітозу, ядерця, мітохондрій. Мають рибосоми клітинною стінкою з пептидогліканом. За типом дихання- аероби та анаероби. Рухаються за допомогою джгутиків, побудованих у прокаріотів з білка флагеліну і не містять мікротрубочок.
Еукаріоти -мають ядро відмежоване від цитоплазми ядерною мембраною, апарат мітозу та ядерця, рибосоми, мітохондрії, не містять пептидоглікану, всі аероби. Рухливість забезпечують джгутики і складаються з білка тубуліна, представляють систему мікротрубочок.
Геном – повний набір генетичної інформації.
Щоб позначити визначення поняття «мікроорганізм» необхідно визначити головний критерій який би відрізняв живе від неживого. Ген є єдиним носієм та зберігачем життя. Відмінність живого від неживого – наявність власної генетичної системи. Усі, хто має свою генетичну систему, розглядаються як організми.
Мікроорганізми - це невидимі простим оком представники всіх царств життя, що займають нижчі щаблі еволюції, але відіграють важливу роль у кругообігу речовин у природі, патології рослин, тварин та людини.
Принципи систематики та класифікації бактерій.
Систематика займається описом видів організмів, з'ясуванням ступеня споріднених відносин між ними та об'єднанням у класифікаційні одиниці (таксони). Класифікація – складова систематики. Вона зводиться до розподілу організмів відповідно до їх ознак за різними таксонами. Таксономія-наука про принципи та методи розподілу (класифікації) організмів в ієрархічному плані. Основною таксономічною одиницею у біології є вид.
Великі таксономічні одиниці: рід, сімейство, порядок, клас. Додаткові категорії: підрід, підтип, підпорядок, підклас.
Вид - група близьких, між собою організмів, що мають загальний корінь походження, на даному етапі еволюції, характеризуються певними морфологічними, біохімічними та фізіологічними ознаками, відокремленим відбором від інших видів та пристосовані до певного середовища проживання.
Специфічні особливості мікроорганізмів, ряд ознак та властивостей використовують для їх класифікації:
1.Морфологічні ознаки - Величина, форма і характер взаєморозташування.
2. Тинкторіальні властивості - здатність фарбуватися різними барвниками. Важлива ознака ставлення до забарвлення за Грамом, яке залежить від структури та хімічного складу клітинної стінки. При руйнуванні клітинної стінки чи втраті (у разі L-трансформації) вони стають грамнегативними. За цією ознакою всі бактерії діляться на грамнегативні (забарвлюються в червоний колір) і грампозитивні (забарвлюються у фіолетовий колір).
3 Культуральні властивості- особливості зростання бактерій на рідких і повних живильних середовищах Зростання на рідких середовищах з утворенням плівки, осаду, помутніння.
Зростання на щільних живильних середовищах у вигляді колоній, можна визначити: форму, розміри, краї колоній, поверхню, прозорість та інші властивості.
У мікробіології використовують спеціальні терміни:
Ø Колонія– видима простим оком ізольована структура, що утворюються внаслідок розмноження та накопичення бактерії за певний термін інкубації.
Ø Термін інкубації- Час зростання бактерій.
Колонія утворюється з однієї батьківської клітини чи кількох ідентичних клітин. Пересівом із ізольованої колонії може бути отримана чиста культура збудника.
Ø Культура- Вся сукупність бактерій, що виросли на щільному або рідкому поживному середовищі.
Ø Чиста культура збудника- один вид бактерій, що виросли на щільному поживному середовищі. Щоб уникнути діагностичних помилок у бактеріології вивчають властивості лише чистих однорідних культур.
Ø Штам- Конкретний зразок цього виду.
4.Рухомість бактерій - Розрізняють рухомі і нерухомі. Рухливі підрозділяються на повзаючі або ковзні, плаваючі, що пересуваються хвилеподібно.
5. Спороутворення - Форма та характер розташування спор у клітці.
6.Фізіологічні властивості - способи харчування, тип дихання, зростання та розмноження.
7.Біохімічні властивості - Здатність ферментувати (розщеплювати) вуглеводи, протеолітична активність, утворення індолу, сірководню.
8.Геносистиматика - Вивчення нуклеотидного складу ДНК та характеристик геному. Точним методом встановлення генетичної спорідненості між бактеріями є визначення ступеня гомології ДНК.Чим більше ідентичних генів, тим вищий ступінь гомології ДНК і ближча генетична спорідненість. Метод молекулярної гібридизації ДНК-ДНКвикористовується для систематики бактерій. Якщо діапазон гомології ДНК від 60 до 100% визначають приналежність одному й тому виду, ступінь гомології від 40 до 60%- до різних пологів.
Морфологія бактерій.
Бактерії мають певну форму і розміри, які виражаються в мікрометрах (мкм). Розрізняють основні форми бактерій: кулясті або кокоподібні (від грец.kokkos-зерно); паличкоподібні, звивисті, ниткоподібні. Крім того існують бактерії, що мають трикутну форму, зіркоподібну, тарілкоподібну, існують бактерії квадратні.
Коккоподібні патогенні бактерії.
Мають форму правильної кулі, деякі бобоподібну та ланцетоподібну. За характером взаєморозташування клітин після поділу коки поділяються на групи:
1. Мікрококи(Від лат. Micros-малий) Діляться в одній площині, розташовуються поодиноко, безладно; сапрофіти, патогенні для людини немає.
2. Диплококи(від лат.diplos - подвійний) Поділ відбувається в одній площині з утворенням пар клітин, що мають або бобоподібну (як представник гонореї- гонокок) Гр (-)-забарвлюються в червоний колір, або ланцетоподібну (як збудник стрептококової пневмонії). Гр (+) – забарвлюються у фіолетовий колір.
3. Стрептококи(Від грец. streptos-ланцюжок) Поділ відбувається в одній площині, але після розмноження клітини зберігають між собою зв'язок, утворюючи ланцюжки різної довжини, що нагадують нитки намиста. Стрептококи є патогенними для людини – викликають ангіну, скарлатину, гнійні запалення та інші. . Гр (+) – забарвлюються у фіолетовий колір.
4. Стафілококи(Від лат. Staphуle - гроно винограду.) Діляться в декількох площинах, утворюючи клітини скупчення, що нагадують гроно винограду. Вони часті збудники гнійних запалень. Гр (+) – забарвлюються у фіолетовий колір.
5. Тетракоки(Від лат. Tetra - чотири.) Розподіл клітин у двох взаємно перпендикулярних площинах з утворенням зошит. Патогенні для людини види зустрічаються рідко. Гр (+) – забарвлюються у фіолетовий колір.
6. Сарцини(від лат.sarcina-зв'язка, тюк.) Розподіл клітин у трьох взаємно перпендикулярних площинах з утворенням пакетів (тюків) з 8, 16, 32 і більшої кількості особин. Зустрічаються у повітрі, існують умовно – патогенні представники.
Гр (+) – забарвлюються у фіолетовий колір.
Паличкоподібні патогенні бактерії.
Термін "Бактерія"(від грец.bakteria - паличка) застосовується як назва всього царства прокаріотів і паличок, що не утворюють суперечку.
Палички, що утворюють суперечки, поділяються на:
Бацили (від лат.bacillus-паличка) представник збудник сибірки (bacillus anthracite).
Клостридії (від латів. clostridium – веретеноподібний) представник збудника правця (Clostridium tetani).
Палички бувають довгими-3мкм як представник збудника газової гангрени (Clostridium novyi); короткими -1,5-3 мкм як кишкова паличка (Escherichia coli) та більшість збудників кишкових інфекцій; дуже короткими-менш 1мкм як представник збудника бруцельозу (brucella meiitensis).
Кінці паличок можуть бути закругленими як у кишкової палички, потовщені як у збудника дифтерії. Паличка може мати яйцеподібну форму як збудник чуми (Yersinia pestis).
По діаметру їх ділять на тонкі як збудник туберкульозу (Mycobactvrium tuberculosis) і товсті як збудник газової гангрени (Clostridium perfringens).
За взаєморозташуванням бактерій їх поділяють на 3 групи:
Монобактерії- палички розташовуються поодиноко і безладно. Гр (-)-фарбуються
Диплобактерії- Розташовуються попарно. у червоний колір
Стрептобацили- Розташовуються ланцюжком.
Звивисті патогенні бактерії.
 За кількістю та характером завитків, а також діаметром клітин поділяють на 2 групи:
За кількістю та характером завитків, а також діаметром клітин поділяють на 2 групи:
 1.Вібріони(від грец.Vibrio- згинаюсь) мають один вигин, представник збудник холери.
1.Вібріони(від грец.Vibrio- згинаюсь) мають один вигин, представник збудник холери.
2. Спірили(Від грец. Speira - завиток) представник спірохети.
Будова бактеріальної клітини.
Клітина – універсальна структурна одиниця всього живого. У її складі можна назвати структури:
I. Клітинна стінка- властиве лише бактеріям (крім мікоплазм). Виконує функції:
Ø Визначає та зберігає постійну форму бактерій.
Ø Захищає клітину від дії механічних та осмотичних сил довкілля.
Ø Бере участь у регуляції росту та поділу клітин.
Ø Забезпечує зв'язок із зовнішнім середовищем.
Клітинна стінка характеризується наявністю унікальної хімічної сполуки-пептидоглікану, що наділяє клітину важливими імунобіологічними властивостями:
Ø Пептидоглікан активізує роботу імунної системи, запускає систему комплементу.
Ø Захищає бактерії особливо грампозитивні від фагоцитозу.
Ø Сприяє розвитку алергічних реакцій. (ГЗТ)
Ø Має протипухлинну дію.
Ø Чинить пірогенну дію на організм тварин і людини.
Ø Порушення його синтезу призводить до перетворення бактерій з S - форми в L - форму за допомогою чого відбувається тривале персистування (знаходження) збудника в організмі - одна з основних причин переходу захворювання з гострої форми на хронічну. Відповідно L – трансформація як і спороутворення є найважливішою формою пристосування бактерій до несприятливих умов існування.
ІІ. Зовнішня мембрана – складається з двох шарів ліпідів та набору білків, локалізованих мозаїчно. Два з основних білків пов'язані з пептидогліканом. Ці білки-поріни утворюють дифузні пори, через які в клітину проникають дрібні молекули. Другорядні білки виконують специфічні функції: одні забезпечують механізми живлення, беруть участь у полегшеній дифузії, інші в активному транспорті молекул через зовнішню мембрану та регуляції клітинного поділу. Зовнішня мембрана виконує функцію бар'єру, через який клітину не здатні проникати великі молекули, що є одним з механізмів стійкості грамнегативних бактерій до антибіотиків.
Якщо бактерії помістити в гіпертонічний розчин, то настає зневоднення клітин, цитоплазма стискається, в результаті клітини гинуть – це явище називається плазмоліз.Цією властивістю використовують для консервування харчових продуктів за допомогою концентрованих розчинів солі або цукру. Стійкі, до плазмолізу збудник ботулізму та золотистий стафілокок, які є частими винуватцям харчових отруєнь.
Якщо бактерії помістити в гіпотонічний розчин або дистильовану воду, відбувається протилежне явище - плазмоптиз-вода спрямовується в клітини, відбувається їх набухання і руйнування.
ІІІ. Цитоплазматична мембрана - є поліфункціональною структурою:
Ø Цитоплазматична мембрана сприймає всю хімічну інформацію, що надходить із зовні.
Ø Осмотичний бар'єр, завдяки чому всередині клітини підтримується певний осмотичний тиск.
Ø Цитоплазматична мембрана разом із клітинною стінкою та бере участь у регуляції росту та клітинного поділу.
Ø Місце генерації енергії у бактерій.
Ø Цитоплазматична мембрана пов'язана зі джгутиками, апаратом регуляції руху.
Ø Цитоплазматична мембрана бере участь у процесах транспорту поживних речовин, у клітину та продуктів життєдіяльності з клітини. У ній містяться білки, що беруть участь у полегшеній дифузії та активному транспорті.
Ø Бере участь у здійсненні біосинтезу білка шляхом стабілізації рибосом.
Ø Бере участь у освіті мезосом.
Цитоплазматична мембрана пов'язана з процесами життєзабезпечення клітини: полегшеною дифузією та активним транспортом. Разом із клітинною стінкою вона утворює оболонку клітини.
IV. Цитоплазма складна колоїдна система. У ній розташовується ядерний апарат – нуклеоплазма, який відокремлений від неї ніякими мембранами. Крім хромосом у цитоплазмі багатьох патогенних бактерій, є плазміди. У цитоплазмі розташовуються рибосоми, мезосоми, макромолекули (тРНК, амінокислоти, нуклеотиди), різні включення (крапельки ліпідів, віск, сірка, гранули глікогену як у клостридій, зерна валютина-як у збудника дифтерії).
V. Периплазматичний простір - знаходиться між цитоплазматичною мембраною та пептидогліканом. Мезосоми та пори з клітинної стінки відкриваються в периплазматичний простір. Цей простір забезпечує взаємозв'язок цитоплазматичної мембрани та клітинної стінки.
VI. Капсула – являє собою слизовий шар, пов'язаний із клітинною стінкою. Вона є зовнішнім покривом бактерії. Деякі патогенні бактерії утворюють капсулу тільки в організмі людини та тварин, як збудники газової гангрени, сибірки. Капсула наділяє бактерію багатьма важливими властивостями:
v утворює оболонку бактерій;
v оберігає від висихання;
v несе запас поживних речовин;
v готують вакцини з компонентів капсули для захисту проти менінгококових та пневмококових інфекцій;
є фактором патогенності для бактерій: вони або маскують їх від фагоцитів, або пригнічують фагоцитоз. Втрата здатності синтезувати капсулу у пневмокока, наприклад, супроводжується повною втратою патогенності.
VII. Джгутики – необхідні руху бактерій. Вони, одержуючи хімічний сигнал із навколишнього середовища, змінюють напрями руху та вибирають оптимальні умови для свого існування. За характером розташування джгутиків та їх кількістю бактерії ділять:
1) Монотрихи- один полярно розташований джгутик (як у холерного вібріона).
2) Лофотріхи-Пучок джгутиків на одному кінці.
3) Амфітріхи- Пучки джгутиків з двох кінців.
4) Перитрихи- безліч джгутиків навколо клітини (як у кишкової палички).
VIII. Спору - захисна форма у несприятливих умовах існування. Це своєрідні клітини, що покоїться. Вони мають високу стійкість до висушування, дії підвищеної температури та хімічних речовин. Високу резистентність (стійкість) суперечку до дії зовнішніх факторів пов'язують із присутністю в оболонці великої кількості Са. Спори в клітині можуть розташовуватися:
v центрально - як у збудника сибірки;
v субтермінально-як у збудника ботулізму;
v термінально – як у збудника правця.
Подібна інформація.
Еукаріоти та прокаріоти. Більшість мікроорганізмів – одноклітинні істоти. Мікробна клітина відокремлена від зовнішнього середовища клітинною стінкою, інколи ж лише цитоплазматичною мембраною і містить різні субклітинні структури. Існують два основних типи клітинної будови, які відрізняються один від одного поруч фундаментальних ознак. Це еукаріотичні та прокаріотичні клітини. Мікроорганізмів, що мають справжнє ядро, називають еукаріот (еу - від грец. Істинний, каріо - ядро). Мікроорганізми з примітивним ядерним апаратом відносяться до прокаріотів (доядерних).
До еукаріотів належать гриби, водорості та найпростіші. За будовою вони подібні до рослинних і тваринних клітин. Бактерії та синьо-зелені водорості (ціанобактерії) відносять до прокаріотів.
В еукаріотичній клітині є ядро, відокремлене від навколишньої цитоплазми двошарової ядерної мембраною з порами. У ядрі знаходяться 1-2 ядерця – центри синтезу рибосомальної РНК та хромосоми – основні носії спадкової інформації, що складаються з ДНК та білка. При розподілі хромосоми розподіляються між дочірніми клітинами внаслідок складних процесів - мітозу та мейозу. Цитоплазма еукаріотів містить мітохондрії, а у фотосинтезуючих організмів - і хлоропласти. Цитоплазматична мембрана, що оточує клітину, перетворюється всередині цитоплазми в ендоплазматичну мережу; є також мембранна органела - апарат Гольджі.
Прокаріотичні клітини влаштовані простіше. Вони немає чіткої межі між ядром і цитоплазмою, відсутня ядерна мембрана. ДНК у цих клітинах не утворює структур, схожих на хромосоми еукаріотів. Тому у прокаріотів не відбуваються процеси мітозу і мейозу. Більшість прокаріотів не утворює внутрішньоклітинних органел, обмежених мембранами.
Крім того, у прокаріотичних клітинах немає мітохондрій та хлоропластів.
Нижче розглядається будова лише прокаріотичної (бактеріальної) клітини, оскільки будова еукаріотичної клітини висвітлено у відповідних курсах ботаніки та зоології.
Форма бактерій. Бактерії, як правило, є одноклітинними організмами, клітина їх має досить просту форму, є кулею або циліндром, іноді вигнутою. Розмножуються бактерії переважно розподілом на дві рівноцінні клітини.

Бактерії кулястої форми називаються коками (лат, coccus - зерно) і можуть бути сферичними, еліпсоїдальними, бобоподібними та ланцетоподібними.
За розташуванням клітин щодо один одного після поділу коки поділяють на кілька форм. Якщо після поділу клітини розходяться і розташовуються поодинці, такі форми називають монококами. Іноді коки при розподілі утворюють скупчень, що нагадують виноградне гроно. Подібні форми відносяться до стафілококів. Коки, що залишаються після поділу в одній площині зв'язаними парами, називаються диплококами, а утворюють різної довжини ланцюжка - стрептококами (рис. 1, 2). Поєднання з чотирьох коків, що з'являються після поділу клітини у двох взаємно перпендикулярних площинах, є тетракоки. Деякі коки діляться у трьох взаємно перпендикулярних площинах, що призводить до утворення своєрідних скупчень кубічної форми, які називаються сардинами.
Більшість бактерій мають циліндричну, або паличкоподібну форму. Раніше всі паличкоподібні форми називалися бацилами (лат. Bacillum - маленька паличка). Після 1875, коли німецький ботанік Ф. Кон відкрив існування суперечка так званої сінної палички, паличкоподібні форми бактерій, що утворюють суперечки, стали називати бацилами, а не утворюють суперечки - бактеріями.
Паличкоподібні бактерії розрізняються за формою, розміром у довжину та в поперечнику, формою кінців клітини, а також за взаємним розташуванням. Вони можуть мати циліндричну форму з прямими кінцями або овальну - із закругленими або загостреними кінцями. Бактерії бувають також злегка вигнутими, зустрічаються ниткоподібні та розгалужені форми (наприклад, мікобактерії та актиноміцети).
Залежно від взаємного розташування окремих клітин після поділу паличкоподібні бактерії ділять на власне палички (поодиноке розташування клітин), диплобактерії або диплобацили (парне розташування клітин), стрептобактерії або стрептобацили (утворюють ланцюжки різної довжини).
Нерідко зустрічаються звивисті, або спіралеподібні, бактерії.
До цієї групи відносяться спірили (від лат. spira - завиток), що мають форму довгих вигнутих (від 4 до 6 витків) паличок, і вібріони (лат. vibrio - згинаюсь), що являють собою лише 1/4 частина витка спіралі, схожі на кому (Рис. 3).
Відомі ниткоподібні форми бактерій, що мешкають у водоймах. Крім перерахованих, зустрічаються багатоклітинні бактерії, що несуть на поверхні клітини протоплазм етичні вирости - простіки, трикутні та зіркоподібні бактерії, а також мають форму замкнутого та незамкнутого кільця та червоподібні бактерії.
Розміри бактерій Клітини бактерій дуже малі. Їх вимірюють у мікрометрах, а деталі тонкої структури – у нанометрах. Коки зазвичай мають діаметр близько 0,5-1,5 мкм. Ширина паличкоподібних (циліндричних) форм бактерій здебільшого коливається від 0,5 до 1 мкм, а довжина дорівнює декільком мікрометрам (2-10). Дрібні палички мають ширину 0,2-0,4 та довжину 0,7-1,5 мкм. Серед бактерій можуть зустрічатися і справжні гіганти, довжина яких сягає десятків і навіть сотень мікрометрів. Форми та розміри бактерій значно змінюються залежно від віку культури, складу середовища та її осмотичних властивостей, температури та інших факторів.
З трьох основних форм бактерій коки найбільш стабільні за розмірами, паличкоподібні бактерії більш мінливі, причому особливо значно змінюється довжина клітин.
Бактеріальна клітина, поміщена на поверхню твердого живильного середовища, росте, ділиться, утворюючи колонію бактерій-нащадків. Через кілька годин зростання колонія складається вже з такої великої кількості клітин, що її можна бачити неозброєним оком. Колонії можуть мати слизову або пастоподібну консистенцію, в деяких випадках вони пігментовані. Іноді зовнішній вигляд колоній настільки характерний, що дозволяє без особливих труднощів провести ідентифікацію мікроорганізмів.
Систематика та номенклатура мікроорганізмів
Численні мікроорганізми (бактерії, гриби, найпростіші, віруси) суворо систематизовані в певному порядку за їх схожістю, відмінностями та взаємовідносинами між собою. Цим займається спеціальна наука, яка називається систематикою мікроорганізмів. Розділ систематики, що вивчає принципи класифікації, називається таксономією.Таксон – група організмів, об'єднана за певними однорідними властивостями у межах тієї чи іншої таксономічної категорії. Найбільшою таксономічною категорією є царство, дрібнішими - підцарство, відділ, клас, порядок, сімейство, рід, вид, підвид та ін.
В основу таксономії мікроорганізмів покладено їх морфологічні, фізіологічні, біохімічні, молекулярно-біологічні властивості. Весь світ мікробів поділяється на три царства:
. царство еукаріотів (гриби та найпростіші);
. царство прокаріотів (бактерії, рикетсії, мікоплазми);
. Царство вірусів.
Еукаріоти подібні до клітин рослин і тварин. Вони мають поверхневу мембрану та внутрішньоклітинну систему елементарних мембран, що становлять ендоплазматичну ретикулярну мережу та комплекс Гольджі. У цитоплазмі еукаріотів міститься оформлене ядро, мітохондрії, рибосоми та ряд інших органел. Розмножуються прості еукаріоти статевим та безстатевим шляхами.
Прокаріоти - організми, що не мають відмежованого ядра, внутрішньоклітинної системи елементарних мембран та мітохондрій, а деякі позбавлені також клітинної стінки. Розмножуються простим поперечним розподілом або брунькуванням.
Однією з основних таксономічних категорій є вид (species) - сукупність особин, що мають загальний корінь походження, подібний генотип та максимально близькі фенотипічні ознаки та властивості.
Сукупність однорідних мікроорганізмів, виділених на живильному середовищі, що характеризується подібними морфологічними, тинкторіальними (ставлення до барвників), культуральними, біохімічними та антигенними властивостями, називається чистою культурою.
Чиста культура мікроорганізмів, виділених із певного джерела і від інших представників виду, називається штамом. Штам - вужче поняття, ніж вид чи підвид. Близьким до штаму є поняття клону; клон - це сукупність нащадків, вирощених із однієї мікробної клітини.
Рішенням Міжнародного конгресу для мікроорганізмів рекомендовано такі таксономічні категорії: царство, відділ, клас, порядок, сімейство, рід, вид.
Назва виду відповідає бінарній номенклатурі, тобто складається із двох слів. Наприклад, кишкова паличка пишеться як Escherichia coli. Перше слово - назва роду, що починається з великої літери, друге слово позначає вигляд і пишеться з малої літери. При повторному написанні виду родова назва скорочується до початкової літери, наприклад, E. Сoli.
Форми бактерій
Всім бактеріям притаманні певні морфологічні властивості (форма, розмір, характер їх розташування в мазку) та тинкторіальні властивості (здатність фарбуватись).Розрізняють 4 основні форми бактерій (рис. 1.1): кулясті (сферичні), або кокоподібні (від грец. kokkos – зерно); паличкоподібні (циліндричні); звивисті (спіралеподібні); ниткоподібні. Крім того, існують бактерії, що мають трикутну, зіркоподібну, тарілкоподібну форму. Виявлено так звані квадратні бактерії, які утворюють скупчення з 8 або 16 клітин у вигляді пласта.
Мал. 1.1. Форми одноклітинних бактерій: а – мікрококи; б - диплококи; в - стрептококи; г - стафілококи; д – сарцини; е - паличкоподібні бактерії; ж - спірили; з - вібріони
Коккоподібні бактерії зазвичай мають форму правильної кулі діаметром 10-15 мкм; деякі - бобоподібну, ланцетоподібну, еліпсоподібну форму. За характером взаєморозташування клітин коки, що утворюються після поділу, поділяють на наступні групи:
1. Мікрококи (від латів. Micros - малий). Клітини діляться в одній площині і найчастіше відразу ж відокремлюються від материнської. Розташовуються поодинці, безладно (рис. 1.1. а).
2. Диплококи (від латів. diplos - подвійний). Розподіл відбувається в одній площині з утворенням пар клітин, що мають або бобоподібну або ланцетоподібну форму (рис. 1.1. б).
3. Стрептококи (від латів. streptos - ланцюжок). Поділ клітин відбувається в одній площині, але клітини, що розмножуються, зберігають між собою зв'язок і утворюють різної довжини ланцюжка, що нагадують нитки бус. Багато стрептококів є шкідливими для людини і викликають різні захворювання: скарлатину, ангіну, гнійні запалення та ін. Наприклад Streptococcus pyogenes (рис. 1.1.в).
4. Стафілококи (від латів. staphyle - гроно винограду). Клітини діляться в декількох площинах, а клітини, що утворюються, розташовуються скупченнями, що нагадують грона винограду (рис. 1.1. г).
5. Тетракоки (від латів. tetra – чотири). Поділ відбувається у двох взаємно перпендикулярних площинах з утворенням зошит.
6. Сарцини (від латів. sarcina – зв'язка, тюк). Поділ відбувається у трьох взаємно перпендикулярних площинах з утворенням пакетів (тюків) з 8-ми, 16-ти, 32-х та більшої кількості особин. Особливо часто зустрічаються у повітрі (рис. 1.1.д).
Паличкоподібні (циліндричні форми) (рис. 1.1.е). За розташуванням палички поділяють:
- на одиночні чи безладно розташовані - монобактерії. Наприклад, Escherihia coli;
- Попарно (по одній лінії) - диплобацили, диплобактерії. Наприклад, Pseudomonas;
- розташовані ланцюжком - стрептобацили, стрептобактерії. Наприклад, Bacillus.
Палички, що утворюють суперечку, поділяють:
- на бацили - аеробні спороутворюючі бактерії. Спору в таких паличок розташовується, як правило, центрально, і її діаметр не перевищує ширини бактерії.
- клостридії – анаеробні спороутворюючі бактерії. Спору вони розташовується термінально чи субтермінально. Вона велика, що розтягує оболонку бактерій, і вони зовні нагадують веретено чи тенісну ракетку.
Звивисті (спіралеподібні) форми
За кількістю та характером завитків, а також за діаметром клітин вони поділяються на три групи:1. Вібріони (від грец. vibrio - звиваюся, згинаюсь) мають один вигин, що не перевищує чверті обороту спіралі. Наприклад, Vibrio (рис. 1.1.з).
2. Спірили (від грецьк. speira - завиток) - клітини, що мають великий діаметр і мале (2-3) кількість завитків. Наприклад - Spirillium minor (рис. 1.1 ж).
3. Спірохети (від грец. speira – завиток, chaita – волосся) – спіралеподібної форми рухливі бактерії.
Ниткоподібні форми
Розрізняють два типи ниткоподібних бактерій: утворюють тимчасові нитки та постійні.Тимчасові нитки (іноді з розгалуженнями) утворюють паличкоподібні бактерії при порушенні умов їх зростання або регуляції клітинного поділу (мікобактерії, коринебактерії, а також рикетсії, мікоплазми, багато грамнегативних та грампозитивних бактерій). При відновленні механізму регуляції поділу та нормальних умов зростання ці бактерії відновлюють звичайні для них розміри.
Постійні ниткоподібні форми утворюються з паличкоподібних клітин, що з'єднуються у довгі ланцюжки або за допомогою слизу, або чохлами, або містками (сіркобактерії, залізобактерії).
Для вивчення тинкторіальних властивостей мікроорганізмів та їхньої морфології використовують анілінові барвники (основні, кислі та нейтральні).
Найбільше застосування мають основні фарби: метиленовий синій, основний фуксин, генціанвіолет, везувін, хризоїдин та ін. Рідше застосовуються нейтральні (нейтральний червоний) та кислі (еозин) фарби. З названих фарб готують спиртові, водно-спиртові та водні розчини. У деяких випадках для підвищення фарбуючої сили розчину до нього додають протрави, наприклад, карболову кислоту, луг та ін.
Для визначення форми бактерій та їх взаємного розташування в мазку використовують прості методи фарбування, тобто фарбування здійснюється одним барвником і мазок виходить забарвленим одним кольором. Наприклад, метиленовий синій. Це фарбування дозволяє краще виявити бобоподібну форму і парне розташування коків.
Для вивчення структури бактеріальної клітини і виявлення особливостей її будови застосовують складні методи забарвлення, які включають цілу низку барвників, протрави і диференціюючі речовини. До складних методів забарвлення належать методи Грама, Нессера, Ожешко та ін.
Л.В. Тимощенка, М.В. Чубик