Цитогенетичні механізми запліднення. Запліднення, його фази, біологічна суть. Як відбувається запліднення яйцеклітини
Запліднення – це складний механізм послідовних процесів, лише за суворого дотримання умов якого можна отримати бажану вагітність. Так, жінка повинна мати дозрілу яйцеклітину, маткові труби, що проходять, достатню товщину слизової оболонки матки, яка повинна бути готова до прикріплення заплідненої яйцеклітини. Після імплантації жіночий організм має забезпечити гормональну підтримку вагітності.
У чоловіка має бути достатня кількість морфологічно нормальних рухливих зрілих сперматозоїдів, здатних через шийку матки проникнути в порожнину матки, маткову трубу та запліднити яйцеклітину.
У жінок статеві клітини називаються яйцеклітини, а у чоловіків – сперматозоїди.
Яйцеклітина нагадує звичайну клітину, має округлу форму, включає ядро, що має у своєму складі ДНК (материнський генетичний матеріал), цитоплазму та оболонку. Клітина оточена так званим «променистим вінцем», що складається із захисних клітин.
Сперматозоїд має принципово іншу будову. Це зумовлено виконуваною ним функцією: йому необхідно подолати відстань від шийки матки до яєчникового відділу маткової труби. Сперматозоїд має головку (у якій міститься ДНК: батьківський генетичний матеріал), шийку та хвостик. На головці у сперматозоїда є спеціальна структура - акросома, що містить ферменти, що сприяють проникненню сперматозоїда в яйцеклітину.
Принципова відмінність жіночого та чоловічого організмів полягає в тому, що запас яйцеклітин у жіночому організмі суворо обмежений та закладається під час внутрішньоутробного розвитку дівчинки. Коли вроджений запас фолікулів виснажується, жінка втрачає репродуктивну функцію (менопауза). У чоловічому організмі процес утворення сперматозоїдів не залежить від віку та носить постійний характер.
Фізіологічний процес запліднення
Процес запліднення залежить від гормональної регуляції, психоемоційного стану жіночого організму, чинників довкілля та інших. Менструальний цикл умовно починається з першого дня менструації, коли відбувається «обнулення» гормонального фону. Саме тоді починається зростання когорти фолікулів, у тому числі до 5-7 дня менструального циклу відбувається відбір одного фолікула. Після досягнення фолікулом розмірів 18-25 мм (зазвичай на 12-14 день менструального циклу) відбувається овуляція.
В результаті відбувається вихід яйцеклітини в черевну порожнину, звідки за допомогою фімбрій (особливих виростів маткової труби) потрапляє до яєчникового відділу маткової труби, де відбувається її зустріч зі сперматозоїдами, які, у свою чергу, після еякуляції пройшли довгий шлях протягом кількох годин жіночим. статевих шляхів. Внаслідок цієї зустрічі, за наявності сприятливих умов середовища, відбувається запліднення.
Надалі, що ділиться ембріон (зігота - містить материнський і батьківський генетичний матеріал) здійснює шлях по маткових труб до матки за рахунок перистальтичних скорочень стінок маткових труб, руху ворсинок і капілярного струму рідини. Надалі, потрапляючи в порожнину матки, ембріон прикріплюється до однієї стінки матки, відбувається імплантація і подальший розвиток ембріона.
Умови, необхідні для успішного процесу запліднення:
дозрівання фолікула в яєчнику, готова до запліднення яйцеклітини;
овуляція; достатня функція жовтого тіла;
нормальна концентрація якісних активних сперматозоїдів;
проникнення сперматозоїдів у маткові труби, їх просування жіночими статевими шляхами, здатність до запліднення;
Запліднення, вихідний момент виникнення нової генетичної індивідуальності, є процесом з'єднання жіночої та чоловічої гамет.
Внаслідок запліднення виникає одноклітинний зародок з диплоїдним набором хромосом та активується ланцюг подій, що лежать в основі розвитку організму.
Біологічне значення запліднення величезне: будучи причиною розвитку нової індивідуальності, воно водночас є умовою продовження життя та еволюції образу.
Слід підкреслити, що запліднення є не одномоментним актом, але саме процес, що займає більш менш тривалий відрізок часу. Це багатоступінчастий процес, у якому розрізняються такі етапи: залучення сперматозоїда яйцем, зв'язування гамет і, нарешті, злиття чоловічих та жіночих статевих клітин. У науковій літературі події, пов'язані зі зближенням гамет іноді називають запліднення розрізняючи зовнішнє і внутрішнє запліднення, залежно від того, виводяться чоловічі статеві клітини в навколишнє середовище або статеві органи жіночої особини. Зовнішнє запліднення характерне для тварин, що мешкають у водному середовищі. Внутрішнє запліднення притаманне головним чином наземним тваринам, хоча воно досить часто зустрічається і у мешканців водного середовища. Осіменіння може бути вільним, при якому всі області ооциту доступні сперміям, але може бути і обмеженим, коли на поверхні яйцеклітини є щільна оболонка з мікропилом. При внутрішньому осіменінні у ряду тварин чоловічі гамети передаються самкам у вигляді сперматофорів, особливих капсул, що містять сперматозоїди Сперматофори спочатку виводяться в навколишнє середовище, а потім у той чи інший спосіб переносяться в статеві шляхи самки.
З'єднання гамет визначає можливість каріогамії, або злиття ядер. Завдяки каріогамії відбувається поєднання батьківських та материнських хромосом, що веде до утворення геному нової особини. В результаті злиття гамет виникає диплоїдна зигота, відновлюється здатність до реплікації ДНК і починається підготовка до поділу дроблення. Механізми активації яйця до розвитку щодо автономні. Їх включення може бути здійснено і крім запліднення, що відбувається, наприклад, при природному чи штучному незайманому розвитку, або партеногенез.
Інтерес до проблеми запліднення виходить далеко за межі власне ембріології. Злиття гамет - плідна модель для вивчення тонких молекулярних і клітинних механізмів специфічної взаємодії клітинних мембран; вивчення молекулярних основ активації метаболізму і проліферації соматичних клітин. Загальнобіологічний інтерес представляє і те, що запліднення є яскравим і, можливо, унікальним прикладом повного обігу клітинної диференціації. Дійсно, високоспеціалізовані статеві клітини не здатні до самовідтворення. Вони гаплоїдні та не можуть ділитися. Однак після злиття вони перетворюються на тотипотентну клітину, яка є джерелом формування всіх клітинних типів, властивих даному організму.
Історія відкриття запліднення губиться у глибині століть. У всякому разі, у XVIII столітті італійський дослідник природи абат Лаццаро Спалланцані (1729-1799) експериментально довів, що запліднення залежить від наявності сперми, і вперше здійснив штучне запліднення яєць жаби, змішуючи їх зі спермою, отриманою з насіння. Проте сенс подій, що при цьому відбувалися, залишався незрозумілим практично до останньої чверті XIX століття, коли Оскар Гертвіг (1849-1922) наприкінці 1870-х років, вивчаючи запліднення у морських їжаків, дійшов висновку, що сутність цього процесу полягає в злитті ядер статевих клітин. Разом з роботами бельгійця Едуарда ван Бенедена (1883, аскарида), німецького вченого Теодора Бовері (1887, аскарида), швейцарського зоолога Германа Фоля (1887, морська зірка) дослідження О. Гертвіга заклали основу сучасних уявлень про. Слід наголосити, що саме ці роботи послужили вагомою підставою для припущення про те, що ядро є носієм спадкових властивостей. Саме Т. Бовері (1862-1915) у серії блискучих цитологічних досліджень обґрунтував наприкінці 1880-х років теорію індивідуальності хромосом та створив основу цитогенетики.
Незабаром після з'ясування сутності запліднення дослідники зосередили увагу на механізмах, що лежать в основі цього процесу. Ця сфера досліджень зберігає актуальність і в наш час. Пальма першості у побудові теорії запліднення належить американському досліднику Франку Ліллі (1862–1915). Вивчаючи властивості «яєчної води», тобто морської води, в якій деякий час знаходилися незапліднені яйця морського їжака Arbacia або поліхети Nereis, Ліллі виявив, що з яєць виділяється речовина, яка має здатність склеювати спермін у грудки. Аглютинація, що спостерігається, виявилася видоспецифічною, і Ліллі назвав фактор аглютинації, що виділяється незаплідненим яйцем, речовиною запліднення, або фертилізином(Від англ. Fertilization – запліднення). Суть висунутої Ліллі теорії запліднення полягає у визнанні того, що в периферичній ділянці яйця знаходиться фертилізин, який має спорідненість до поверхневих рецепторів спермію (антифертилізин спермію). Завдяки цій спорідненості фертилізин пов'язує, згідно з Ліллі, спермії. Однак, щоб претендувати на універсальність і пояснити не тільки механізм з'єднання гамет, а й причини аглютинації сперміїв, можливість запобігання поліспермії, високу специфічність процесу запліднення і т.д., теорія фертилізину потребувала численних припущень, під гнітом яких вона врешті-решт і.
Вже під час ранніх досліджень запліднення виникло уявлення про гамони - речовини, які забезпечують активацію чи блокування окремих етапів запліднення. Відповідно до їх походження розрізняли гіногамони, що виділяються яйцеклітинами, і андрогамони, що виробляються чоловічими статевими клітинами. Так, вважали, що гіногамон 1, дифузуючи з яйця, активує рух сперматозоїда, долаючи дію андрогамону 1, який пригнічує рух сперматозоїда. Гіногамон 2 – синонім фертилізину, а андрогамон 2 – антифертилізину спермію.
У п'ятдесяті роки XX століття ідея взаємодії фертилізину з антифертилізином трансформувалася на гіпотезу специфічного фагоцитозу. Згідно з цією концепцією, наявність на поверхні яйця і спермія молекул, що взаємодіють, забезпечує комплементарну реакцію за принципом застібки «блискавки», завдяки якій спермій виявляється поглиненим яйцем.
Незважаючи на відому умоглядність, ці та багато інших подібних гіпотез про механізми взаємодії сперматозоїдів і яєць відіграли свою позитивну роль, виявивши, по-перше, існування цілого сімейства специфічних молекул на поверхні взаємодіючих гамет і, по-друге, започаткувавши планомірні дослідження природи цих молекул .
Друга половина минулого століття – період розквіту ультраструктурних та молекулярно-біологічних досліджень, які виявили велику різноманітність конкретних форм клітинної взаємодії при заплідненні. Стало ясно, що універсальна теорія запліднення, якщо й може існувати, лише як зведення деяких найзагальніших принципів організації цього процесу.
Конкретні механізми запліднення залежать від багатьох чинників. Досить сказати про своєрідність запліднення у тварин із зовнішнім та внутрішнім заплідненням. Очевидно, що певні відмінності процесу запліднення обумовлені і тим, що у різних тварин проникнення спермію в яйце відбувається на різних етапах оогенезу. У багатьох анелід, молюсків, нематод і ракоподібних сперматозоїд проникає в ооцити першого порядку на стадії профази. В інших кільчастих черв'яків, молюсків та у комах – на стадії метафази первинного ооциту. Для багатьох хребетних характерне запліднення на стадії метафази вторинного ооциту. У деяких кишковопорожнинних та у морських їжаків запліднення відбувається на стадії зрілого яйця вже після завершення поділів дозрівання та виділення направних, або редукційних тілець. Нарешті, не можна не згадати і різноманітність типів сперматозоїдів, серед яких є джгутикові форми та спермін без джгутиків (наприклад, амебоїдні спермін нематод), з акросомою та без неї, що мають акросомну нитку та позбавлені її. Природно, що в такому разі конкретні механізми, що забезпечують тонку взаємодію між статевими клітинами, різняться.
Якщо ви знайшли помилку, будь ласка, виділіть фрагмент тексту та натисніть Ctrl+Enter.
Як відомо, після досягнення статевої зрілості у кожної дівчини, а потім жінки один раз на місяць відбувається. Це досить складний фізіологічний процес, в ході якого зріла яйцеклітина виходить з яєчника в маткову трубу. Саме у ній і відбувається запліднення.
Особливості овуляції
Злиття сперматозоїдів з яйцеклітиною відбувається протягом дванадцятої години після того, як вона виходить у маткову трубу. Час овуляції розрахувати нескладно, і з найдостовірніших методів визначення є , тобто температури у прямій кишці. Ця процедура має щоденно проводитися протягом декількох місяців. Температура вимірюється в той самий час, рано вранці, не встаючи в ліжку, за допомогою звичайного градусника.
Якщо занести дані до графіка, можна побачити криву дозрівання своєї яйцеклітини. Перед початком менструації температура максимально знижується, а момент овуляції настає або в останній день зниженої температури, або в перший день її підвищення. Найсприятливішим днем для запліднення яйцеклітини є той, коли настає овуляція або кілька днів до її початку.
Це тим, що сперматозоїди, які потрапили в порожнину маткової труби, залишаються життєздатними протягом кількох діб. Знаючи день настання овуляції, можна не тільки зачати дитину, але й спробувати. Для цього існують різні календарі і зачаття.
Механізм запліднення
Запліднення яйцеклітини є тривалим і складним механізмом, під час якого відбувається поєднання чоловічої та жіночої статевих клітин. Насіннєва рідина, яка під час статевого акту потрапляє до жіночої піхви, містить приблизно від 60 до 150 млн. зрілих сперматозоїдів. За рахунок того безперервного скорочення матки, насіннєва рідина їй активно захоплюється, у зв'язку з чим рухливі сперматозоїди просуваються в порожнину матки протягом декількох хвилин, а потім досягають далеких відділів маткової труби, де розташовується яйцеклітина.
Незважаючи на те, що чоловічих статевих клітин багато, вони зустрічають безліч перешкод на своєму шляху (кисле середовище піхви, слизовий вміст каналу шийки матки і так далі), і тільки один найшвидший сперматозоїд зможе запліднити яйцеклітину. Щоправда, численними дослідженнями доведено, що у яйцеклітину можуть проникнути кілька сперматозоїдів, але ядро зі спадковою інформацією яйцеклітини може з'єднатися з ядром лише одного сперматозоїда, у результаті утворюється лише один ембріон. Звичайно, бувають випадки, коли в процесі запліднення виходить кілька ембріонів, і народжуються близнюки.
Сперматозоїд проходить через міцні оболонки жіночої клітини завдяки розчиненню ферментами, що містяться в акросомній капсулі головки. Вступаючи з контакт з яйцеклітиною, капсула розривається, і з неї до оболонок починає прикріплюватися акросомна нитка та виділятися речовини, що руйнують оболонку яйцеклітини. Розчинивши невелику ділянку, акросомна нитка проникає вглиб яйцеклітини та щільно з'єднається з її внутрішнім вмістом. Потім ядро та внутрішній вміст головки сперматозоїда всмоктується всередину жіночої статевої клітини.
Зміни у яйцеклітині
Повне проникнення сперматозоїда у жіночу статеву клітину запускає процес суттєвої зміни фізіологічних процесів у ній. Оболонки яйцеклітини стають набагато більш проникними, що дуже важливо для активного накопичення поживних речовин, за допомогою яких розвиватиметься ембріон. Починають більш активно вироблятися білки, кальцій та вуглеводи, вбирається максимальна кількість кальцію та фосфору – загалом, ведеться підготовка до розвитку плода.
Найбільш важливі та значні для майбутньої дитини події відбуваються протягом приблизно дванадцятої години після проникнення сперматозоїда в яйцеклітину. У цей час ядра чоловічої та жіночої клітин, що несуть у собі всю спадкову інформацію, з'єднуються. Утворюється нова клітина з повним набором хромосом, з якої потім розвинеться ембріон і в результаті народиться нова людина.
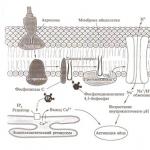
Після проникнення в статеві шляхи самки, сперматозоїд виявляє запліднюючу здатність після процесу Капоцитації. Його суть: головка сперматозоїда має ділянки, що містять фермент глікозилтрансферазу. Але цей фермент блокований галактозом та N-ацетилглюкозаміном. Глікопротеїди, що виділяються у статеві шляхи самки, звільняють блокуючі ферменти. Тоді сперматозоїд здатний пізнавати N-ацетилглюкозамінові залишки в зоні пілюциду (оболонка, вкрита шаром фолікулярних клітин). Тоді фермент знаходить потенційний субстрат. Далі йде 2-й процес, який ініціюється оболонками яйцеклітини. Акросомальнареакція. Її механізм: після контакту зі драглистим оболонкою, в сперматозоїд надходять іони Са. При зовнішньому заплідненні іони Са надходять із води, а при внутрішньому з ендоплазматичного ретикулюму. Паралельно йде процес перебудови мембранних процесів, що забезпечують надходження всередину Na і протонів поза. Так йде підвищення рН, що веде до полімеризації актину. Далі активізується домінова АТФаза. Потім відбувається екзоцитоз акросомальної бульбашки – подвійна мембрана замінюється на одинарну. На акросомальному вирості, що утворився, з'являється білок бендин (дізнається рецептори на яйцеклітині). У хвостатих амфібій, рептилій і птахів у яйце досить часто проникає не один, а кілька сперматозоїдів, і у яєць цих тварин виробилися спеціальні механізми, що інактивують ядра надмірних сперматозоїдів. Більшість інших хребетних ПолісперміяЗапобігається поверхневим реакціям, які перешкоджають проникненню в яйце більш ніж одного сперматозоїда. У яйцях таких тварин є поверхневий шар кортикальних гранул; у яйцях тих хребетних, які допускають проникнення кількох сперматозоїдів, таких гранул немає. У хребетних, що допускають проникнення в яйце лише одного сперматозоїда, перша реакція, що виникає у відповідь на злиття сперматозоїда з яйцем, полягає у швидкій зміні електричних властивостей плазматичної мембрани яйця. Позитивний мембранний потенціал перешкоджає виникненню поліспермії, тоді як зниження потенціалу у щойно заплідненого яйця робить її можливою. Акросома сперматозоїда містить гідро- та ротелітичні ферменти, наприклад, акрозин, подібний до хемотрипсину. Акросома містить фермент (георуронідаза), що розщеплює променистий вінець. Ці ферменти забезпечують проникнення сперматозоїда в яйцеклітину. У зоні контакту відбувається дезантеграція мембрани яйцеклітини та сперматозоїда. У зоні контакту утворюються міцели з утворенням пролому та вміст сперматозоїду проникає всередину. Подія, що перешкоджає поліспермії і виникає через кілька хвилин після проникнення в яйце сперматозоїда, є Кортикальна реакція.Кортикальні гранули, починаючи з тієї точки, в якій відбулося злиття яйця зі сперматозоїдом, переміщуються до внутрішньої поверхні плазматичної мембрани, зливаються з нею, а потім виділяють свій вміст у простір, що оточує яйце. Після вивільнення вмісту кортикальних гранул проникнення в яйце інших сперматозоїдів блокується змінами в зоні пілюциду та плазматичної мембрани яйця. Механізм корт. реак. схожий на акросомальну реакцію – екзоцитоз ферментів у простір між плазматичною мембраною та жовтковою оболонкою. У цих гранулах є полісахариди, які забезпечують проникнення води. Крім води потрапляють інші речовини. Гіолін створює на мембрані гіоліновий шар, що забезпечує утримання бластомерів при дробленні. Ще утворює захист від сперматозоїдо. У ссавців є реакція зони, коли сперматозоїди проникають усередину, на яйцеклітині змінюються рецептори та не дають проникненню іншим сперматозоїдам. Після проникнення сперматозоїда в яйцеклітину відбувається деконденсація генетичного матеріалу та руйнування ядерної оболонки. Навколо деконденсованого генетичного матеріалу утворюється нова оболонка. Утворюється 2 пронуклюуси, що здійснюють рухи - танець пронуклюксів. Після цього оболонки ядер дезонтегруються і хромосоми подвоюються з наступним мітотичним поділом. Це останній етап, що має перешкоду для гібридизації.
Механізми запліднення
Процес запліднення у тварин можна поділити на три фази. Перша фаза характеризується зближенням сперматозоїда з яйцеклітиною до контакту. У цю фазу здійснюються дистантні взаємодії між статевими клітинами. Друга фаза починається з того, що сперматозоїд прикріплюється до поверхні яйцеклітини. У цей час спостерігаються контактні взаємодії між статевими клітинами. Третя фаза процесу запліднення починається після проникнення сперматозоїда в яйце і завершується об'єднанням ядер чоловічої та жіночої статевих клітин. Ця фаза характеризує взаємодію всередині яйця.
Дистантні взаємодії між статевими клітинами
Дистантні взаємодії забезпечуються низкою неспецифічних факторів, серед яких особливе місце належить хімічним речовинам, що виробляються статевими клітинами. Відомо, що статеві клітини виділяють гамони чи гормони гамет. Гамони, що виробляються яйцеклітинами, називають гіногамонами, а спрематозоїдами – андрогамонами. Жіночі статеві клітини виділяють дві групи гамонів: гіногамони I та гіногамони II, що впливають на фізіологію чоловічих статевих клітин. Сперматозоїди виробляють андрогамони І та ІІ.
Деякі з цих хімічних речовин спрямовані на підвищення ймовірності зустрічі сперматозоїда з яйцеклітиною. Відомо, що рух сперматозоїда до яйця здійснюється через посередництво хемотаксису - рух сперматозоїдів за градієнтом концентрації деяких хімічних речовин, що виділяються яйцеклітиною. Хемотаксис достовірно показаний для багатьох груп тварин, особливо безхребетних: молюсків, голкошкірих та напівхордових. Хемотактичні фактори виділені з яйцеклітин морських їжаків: в одних видів - це пептид, що складається з десяти амінокислот, і названий сперактом, в інших видів - пептид складається з чотирнадцяти амінокислот і отримав назву резакту. При додаванні екстрактів цих речовин у морську воду сперматозоїди відповідного виду починають рухатися вгору по градієнту їх концентрації.
У русі сперматозоїдів ссавців по верхніх відділах яйцеводи істотне значення має явище реотаксису - здатність рухатися проти зустрічної течії рідини яйцеводи.
Після того, як сперматозоїд пройде крізь захисні оболонки яйця та вступить у контакт з його плазматичною мембраною, починаються контактні взаємодії між статевими клітинами, які призведуть до проникнення сперматозоїду до цитоплазми яйця.
Контактні взаємодії між статевими клітинами
Контакт сперматозоїда з мембраною яйцеклітини призводить до активації статевих клітин. Реакція активації пов'язана зі складними морфологічними, біохімічними та фізико-хімічними змінами у статевих клітинах. Активація чоловічої статевої клітини, в першу чергу, пов'язана з акросомною реакцією, а жіночої - з кортикальною реакцією.
Акросомна реакціяхарактеризується швидкими змінами в акросомному апараті головки сперматозоїда, що супроводжуються вивільненням ув'язнених у ній спермолізинів та викиданням акросомної нитки у бік поверхні яйця.
Розглянемо загальну схему акросомної реакції у представників різних груп морських безхребетних - голкошкірих, кільчастих черв'яків, двостулкових молюсків, кишково-дихаючих та ін.
На вершині головки сперматозоїда, плазматична мембрана і частина мембрани акросомного бульбашки, що прилягає до неї, розчиняються (лізуються). Вільні краї обох мембран зливаються між собою єдину мембрану. З акросоми, що оголилася, виходять спермолізини в навколишнє середовище і призводять до розчинення яйцевих оболонок у місці контакту зі сперматозоїдом. Після цього внутрішня мембрана акросмного апарату випинається назовні і утворює виріст у вигляді трубочки (акросомна нитка). Акросомна нитка подовжується, проходить через розпушену область додаткових яйцевих оболонок і вступає в контакт із плазматичною мембраною яйцеклітини. В ділянці контакту акросомної нитки з поверхнею яйця плазматичні мембрани зливаються і вміст акросомної трубочки (нитки) з'єднується з цитоплазмою яйцеклітини. В результаті злиття мембран утворюється цитоплазматичний місток. Трохи пізніше цитоплазматичним містком в цитоплазму яйця перейдуть ядро і центріоль сперматозоїда. Акросомна реакція завершується вбудовуванням мембрани сперматозоїда в мембрану яйцеклітини. З цього моменту сперматозоїд та яйцеклітина є вже єдиною клітиною (Рис.7, 8, 9.).
Рис.7. Акросомна реакція сперматозоїда:А - В - злиття зовнішньої мембрани акросоми та мембрани сперматозоїда. Виливання вмісту акросомної бульбашки; 1 – мембрана акросоми; 2 - мембрана сперматозоїда; 3 – глобулярний актин; 4 – ферменти акросоми; Г - Д - полімеризація актину та утворення акросомного виросту; 5 - біндін; 6 – виріст акросоми; 7 – актинові мікрофіломенти; 8 – ядро сперматозоїда. (за Голіченковим)
При загальній схожості акросомної реакції у цих тварин між ними є і певні відмінності. Так, у голкошкірих на відміну у хробаків та молюсків в акросомному апараті не містяться літичні ферменти. У більшості вивчених тварин утворюється одна акросомна нитка, а в деяких черв'яків – кілька таких ниток.

Рис.8. Послідовність акросомної реакції у морського їжака.(за Голіченковим)
При заплідненні хребетних тварин також відбувається акросомна реакція. У нижчих хребетних (міноги, і осетрові риби), вона багато в чому подібна до акрсомной реакцією сперміїв безхребетних тварин.

Рис.9. Схема процесів, що відбуваються при взаємодії мембран яйцеклітини та сперматозоїда в ході запліднення (за Гілбертом).
У акулових риб, рептилій і птахів, яйця яких одягнені щільними оболонками, з'єднання гамет відбувається раніше, ніж сформуються. У цих тварин акросома продовжує виконувати свою початкову роль і добре розвинена.
Акросомна реакція у ссавців відрізняється від такої реакції у хребетних та нижчих хребетних. У спермії ссавців акросомна реакція протікає без утворення акросомного виросту, наблизившись до поверхні яйця, спермій зливається з його плазматичною мембраною бічною поверхнею головки.
У комах і вищих риб з'єднання статевих клітин відбувається після того, як повністю утворюються додаткові щільні яйцеві оболонки. У цих випадках сперматозоїд проникає в яйце через мікропілярні канали та з'єднання гамет відбувається без участі акросоми.
Активація яєць. Кортикальна реакція.Після того, як чоловіча статева клітина прикріпиться до поверхні яйця та її акросомна нитка вступить у контакт із поверхнею ооплазми, відбувається активація яйцеклітини. Активація яйця пов'язана зі складними змінами різних сторін його діяльності. Найбільш яскравим зовнішнім проявом активації є зміни поверхневого шару ооплазми, що отримали назву кортикальної реакції (рис. 10).

Рис.10. Кортикальна реакція в яйці морського їжакаА-наближення спермію до яйця; Б-Г-послідовні стадії кортикальної реакції; показано хвилю виділення вмісту кортикальних гранул, що поширюється від місця проникнення спермію, відділення оболонки та утворення перивітеллінового простору, формування гіалтнового шару; гс-гіаліновий шар; жовткова об болочка кг-кортикальна гранула; оо-оболонка запліднення пм-плазматична мембрана; пп-перивітеліновий простір, заповнений перивітеліновою рідиною (по Гінзбург).
Розглянемо послідовні стадії кортикальної реакції з прикладу найповніше, вивчених яйцеклітин морського їжака. Кортикальна реакція починається з того, що мембрана, що обмежує кожну кортикальну гранулу, злипається із плазматичною мембраною яйця. Тут гранули відкриваються, і їх вміст виливається в жовткову оболонку. Процес секреції вмісту кортикальних гранул починається від місця прикріплення сперматозоїда і хвилеподібно поширюється на всі боки до тих пір, поки не охопить всю поверхню яйця. Частина виділеного вмісту кортикальних гранул одержується і розчиняється, утворюючи перивітеллінову рідину, яка відтісняє жовткову оболонку від плазмолеми яйця, що призводить до збільшення об'єму перивітелінового простору. Інша частина вмісту кортикальних гранул зливається з жовтковою оболонкою, яка при цьому потовщується та перетворюється на оболонку запліднення. Частина кортикальних гранул, що не беруть участі в утворенні оболонки запліднення, перетворюються на щільний шар, званий гіаліновим шаром, розташованим над плазматичною мембраною. Після того, як сформується оболонка запліднення, інші сперматозоїди втрачають можливість проникнути в ооплазму яйця.
В останні роки було вивчено хімічний склад вмісту кортикальних гранул. Показано, що вміст кортикальних гранул містить наступні речовини: а) протеолітичний фермент (актелінова справаміназа), що розриває зв'язки між клітинною оболонкою та плазматичною мембраною яйця; б) протеолетичний фермент (сперм-рецепторна гідролаза), який звільняє сперму, що осіла на жовтковій оболонці; в) глікопротеїд, що втягує воду в простір між жовтковою оболонкою та плазматичною мембраною, викликаючи їхнє розшарування; г) фактор, що сприяє утворенню оболонки запліднення; д) структурний білок гіалін, що бере участь у освіті гіалінового шару.
Яке біологічне значення кортикальної реакції?
По-перше, кортикальна реакція є тим механізмом, який захищає яйце від проникнення надчислених сперматозоїдів.
По-друге, що утворюється в результаті кортикальної реакції перивітелінова рідина, служить специфічним середовищем, в якому протікає розвиток зародка.
При активації яйця спостерігаються й інші зміни різних сторін його діяльності.
По-перше, знижується гальмо, яке блокувало мейоз і, ядерні перетворення тривають з тієї самої стадії, на якій вони зупинилися до моменту виходу яйця з яєчника.
По-друге, спостерігається серія біохімічних змін, що супроводжуються посиленням вуглеводного обміну, підвищенням синтезу ліпідів та білків.
По-третє, різко зростає проникність клітинної мембрани для іонів натрію та калію.
Події, що відбуваються в яйці після проникнення сперматозоїда
Після того, як плазматична мембрана акросомної нитки спермію зливається з плазматичною мембраною яйця, спермій втрачає свою рухливість і його залучення всередину яйця відбувається завдяки дії сил, що виходять з активованого яйця. Зазвичай сперматозоїд втягується в ооплазму разом із хвостом, але іноді хвостовий відділ відкидається. Однак і в тих випадках, коли джгутик проникає в яйце, він відкидається та розсмоктується.
Висококонденсоване ядро сперматозоїда починає набухати, хроматин розпушується і ядро перетворюється на своєрідну структуру, що називається чоловічим пронуклеусом.
Аналогічні зміни відбуваються й у ядрі яйцеклітини, у результаті утворюється жіночий пронуклеус. У період формування пронуклеусів, вздовж хромосом відбувається реплікація ДНК. Надалі пронуклеуси починають переміщатися до центру яйцеклітини. Ядерні оболонки, що оточують кожного з пронуклеусів, руйнуються, пронуклеуси зближуються і відбувається каріогамія. Каріогамія – це остання стадія запліднення. При поєднанні пронуклеусів утворюється ядро з набором диплоїдним хромосом. Потім хромосоми займають екваторіальне положення, і настає перший поділ зиготи.
Ооплазматична сегрегація.Після проникнення сперматозоїда починаються інтенсивні переміщення цитоплазми яйцеклітини (ооплазми). У цьому відбувається розшарування, відмішування різних складових частин ооплазми, що позначається як ооплазматическая сегрегація. У процесі цього намічаються основні елементи просторової організації зародка. Тому даний етап розвитку називають також проморфогенез: мається на увазі, що в цей час хіба що розставляються віхи для майбутніх морфогенетичних процесів.
Моно- та поліспермія
Проникнення в яйцеклітину одного сперматозоїда отримало назву фізіологічної моноспермії. Моноспермія властива всім групам тварин із зовнішнім заплідненням і багатьом тваринам із внутрішнім заплідненням (тим, які подібно, ссавцям мають яйцеклітини невеликого розміру).
В інших тварин, наприклад, у деяких членистоногих (комахи), молюсків (клас черевоногих), хордових (акулоподібні риби, хвостаті амфібії, рептилії та птиці) в яйцеклітину проникає велика кількість сперматозоїдів. Таке явище отримало назву фізіологічної поліспермії. Однак і в цьому випадку з ядром яйцеклітини з'єднується лише ядро одного сперматозоїда, тоді як інші руйнуються (рис.11).

Мал. 11. Поліспермія у тритону.А-проникнення сперміїв у яйце на стадії метафази II поділу дозрівання; Б-синхронні зміни насіннєвих ядер, утворення насіннєвих зірок; В-жіноче ядро з'єднується з одним із насіннєвих ядер; Г - Е-синкаріон вступає в мітоз, надчислені насіннєві ядра відтісняються у вегетативну півкулю і дегенерують. Цифри над зображенням яєць - час після проникнення сперміїв при температурі 23 о (Гінзбург).
При фізіологічній моноспермії є спеціальні механізми захисту яйця від поліспермії. Перший механізм пов'язаний із зміною мембранного потенціалу. Встановлено, що в яйцеклітині жаби через кілька секунд після контакту зі сперматозоїдом заряд мембрани змінюється від -28 до 8 мв і залишається позитивним протягом 20 хв. Такі ж зміни мембранного потенціалу було виявлено в яйцеклітинах морського їжака. Виявилося, що позитивний заряд мембрани перешкоджає поліспермії. Інший широко поширений механізм захисту яйця від проникнення надчислених сперматозоїдів пов'язаний з утворенням оболонки запліднення та перивітелінової рідини.