Viljastamise tsütogeneetilised mehhanismid. Viljastumine, selle faasid, bioloogiline olemus. Kuidas toimub munaraku viljastumine?
Viljastamine on keerukas järjestikuste protsesside mehhanism, mille tingimuste range järgimisel on võimalik saada soovitud rasedus. Seega peab naisel olema küps munarakk, läbitavad munajuhad ja piisava paksusega emaka limaskest, mis peab olema valmis viljastatud munaraku kinnitumiseks. Pärast implanteerimist peab naisorganism pakkuma raseduseks hormonaalset tuge.
Mehel peab olema piisav arv morfoloogiliselt normaalseid liikuvaid küpseid seemnerakke, mis on võimelised läbi emakakaela emakaõõnde, munajuhasse tungima ja munarakku viljastama.
Naistel nimetatakse sugurakke munadeks ja meestel sperma.
Munarakk meenutab tavalist rakku, on ümara kujuga, sisaldab DNA-d (ema geneetiline materjal) sisaldavat tuuma, tsütoplasmat ja membraani. Rakku ümbritseb nn kiirgav kroon, mis koosneb kaitserakkudest.
Sperma on põhimõtteliselt erineva struktuuriga. See on tingitud funktsioonist, mida see täidab: see peab ületama kauguse emakakaelast munajuha munasarjaosani. Spermal on pea (mis sisaldab DNA-d: isa geneetilist materjali), kael ja saba. Sperma peas on spetsiaalne struktuur - akrosoom, mis sisaldab ensüüme, mis hõlbustavad sperma tungimist munarakku.
Põhiline erinevus emas- ja isasorganismide vahel seisneb selles, et naisorganismi munavaru on rangelt piiratud ja see muneb tüdruku sünnieelse arengu ajal. Kui kaasasündinud folliikulite varu on ammendunud, kaotab naine reproduktiivse funktsiooni (menopaus). Meeste kehas ei sõltu sperma moodustumise protsess vanusest ja on pidev.
Viljastamise füsioloogiline protsess
Viljastumise protsess sõltub täielikult hormonaalsest regulatsioonist, naise keha psühho-emotsionaalsest seisundist, keskkonnateguritest jne. Menstruaaltsükkel algab tavapäraselt menstruatsiooni esimesel päeval, mil hormonaalne taust on "nullitud". Siis algab folliikulite kohordi kasv, millest menstruaaltsükli 5.-7. päevaks valitakse välja üks folliikul. Kui folliikuli suurus on 18–25 mm (tavaliselt menstruaaltsükli 12.–14. päeval), toimub ovulatsioon.
Selle tulemusena vabaneb munarakk kõhuõõnde, kust fimbriate (munajuha spetsiaalsed väljakasvud) abil satub see munajuha munasarjaosasse, kus kohtub spermaga, mis omakorda pärast ejakulatsiooni läbib naiste suguelundite kaudu mitu tundi pikka teekonda. Selle kohtumise tulemusena toimub soodsate keskkonnatingimuste olemasolul väetamine.
Seejärel liigub juba jagunev embrüo (sügoot – sisaldab ema ja isa geneetilist materjali) munajuhade kaudu emakasse munajuhade seinte peristaltiliste kontraktsioonide, villi liikumise ja vedeliku kapillaaride voolu tõttu. Seejärel kinnitub embrüo emakaõõnde sisenedes ühe emaka seina külge, toimub implantatsioon ja embrüo edasine areng.
Edukaks viljastamisprotsessiks vajalikud tingimused:
folliikuli küpsemine munasarjas, munarakk on viljastamiseks valmis;
ovulatsioon, kollase keha piisav funktsioon;
kvaliteetse aktiivse sperma normaalne kontsentratsioon;
spermatosoidide tungimine munajuhadesse, nende liikumine läbi naiste suguelundite, võime viljastada;
Viljastumine, uue geneetilise individuaalsuse tekkimise esialgne hetk, on naiste ja meeste sugurakkude ühendamise protsess.
Viljastamise tulemusena tekib diploidse kromosoomikomplektiga üherakuline embrüo ja aktiveerub organismi arengu aluseks olev sündmuste ahel.
Viljastamise bioloogiline tähendus on tohutu: olles eelduseks uue individuaalsuse kujunemisele, on see samal ajal ka elu jätkumise ja liigi evolutsiooni tingimus.
Tuleb rõhutada, et väetamine ei ole ühekordne toiming, vaid pigem protsess, mis võtab rohkem või vähem aega. See on mitmeetapiline protsess, milles eristatakse järgmisi etappe: sperma ligimeelitamine munaga, sugurakkude sidumine ja lõpuks isas- ja naissugurakkude liitmine. Teaduskirjanduses nimetatakse sugurakkude konvergentsiga seotud sündmusi mõnikord ka seemendamiseks, eristades välist ja sisemist viljastumist, olenevalt sellest, kas meeste sugurakud satuvad keskkonda või naiste suguelunditesse. Väline seemendamine on tüüpiline veekeskkonnas elavatele loomadele. Sisemine seemendamine on iseloomulik peamiselt maismaaloomadele, kuigi veekeskkonna elanike seas on see üsna levinud. Seemendamine võib olla tasuta, mille käigus on spermale ligipääsetavad kõik munaraku piirkonnad, kuid võib olla ka piiratud, kui munaraku pinnal on tihe membraan mikropüüliga. Paljude loomade sisemise seemenduse käigus kanduvad isassugurakud emasloomadele vormis spermatofoorid, spetsiaalsed spermat sisaldavad kapslid. Spermatofoorid satuvad esmalt keskkonda ja seejärel ühel või teisel viisil emaslooma suguelunditesse.
Sugurakkude ühendus määrab võimaluse kariogaamia või tuumasünteesi. Tänu kariogaamiale toimub isa ja ema kromosoomide ühinemine, mis viib uue indiviidi genoomi moodustumiseni. Sugurakkude ühinemise tulemusena tekib diploidne sügoot, taastub DNA replikatsioonivõime ja algab ettevalmistus lõhustumiseks. Muna arenguks aktiveerimise mehhanismid on suhteliselt autonoomsed. Nende kaasamine võib toimuda lisaks väetamisele, mis toimub näiteks loodusliku või kunstliku neitsi arengu käigus või partenogenees.
Huvi viljastamise probleemi vastu ületab embrüoloogia enda ulatuse. Gamete fusioon on viljakalt kasutatav mudel spetsiifiliste rakumembraanide interaktsioonide peenmolekulaarsete ja rakuliste mehhanismide uurimiseks; uurida somaatiliste rakkude metaboolse aktivatsiooni ja proliferatsiooni molekulaarseid aluseid. Üldist bioloogilist huvi pakub ka see, et viljastamine on rabav ja võib-olla ainulaadne näide rakkude diferentseerumise täielikust pöördumisest. Tõepoolest, kõrgelt spetsialiseerunud sugurakud ei ole võimelised ise paljunema. Nad on haploidsed ega suuda jaguneda. Pärast sulandumist muutuvad nad aga totipotentseks rakuks, mis toimib kõigi antud organismile omaste rakutüüpide moodustumise allikana.
Viljastamise avastamise ajalugu on aegade hämarusse kadunud. Igatahes tõestas itaalia loodusteadlane abt Lazzaro Spallanzani (1729-1799) 18. sajandil eksperimentaalselt, et viljastumine sõltub sperma olemasolust, ning teostas esmakordselt konnamunade kunstliku viljastamise, segades need kokku seemnerakkudega. munandid. Sellegipoolest jäi sel juhul aset leidnud sündmuste tähendus ebaselgeks peaaegu kuni 19. sajandi viimase veerandini, mil Oscar Hertwig (1849-1922) jõudis 1870. aastate lõpus merisiiliku viljastumist uurides järeldusele, et see protsess on idurakkude tuumade liitmine. Koos belglase Eduard van Benedeni (1883, ümaruss), saksa teadlase Theodor Boveri (1887, ümaruss) ja Šveitsi zooloogi Hermann Fohli (1887, meritäht) töödega panid O. Hertwigi uurimused aluse kaasaegsetele ideedele väetamine. Tuleb rõhutada, et just need tööd olid tugevaks aluseks eeldusele, et tuum on pärilike omaduste kandja. See oli T. Boveri (1862-1915) hiilgava tsütoloogiliste uuringute seerias, kes 1880. aastate lõpus põhjendas kromosoomi individuaalsuse teooriat ja lõi tsütogeneetika aluse.
Varsti pärast viljastamise olemuse selgitamist keskendusid teadlased oma tähelepanu selle protsessi aluseks olevatele mehhanismidele. See uurimisvaldkond on tänapäeval aktuaalne. Viljastamise teooria väljatöötamise juhtroll kuulub Ameerika teadlasele Frank Lillyle (1862-1915). Uurides “munavee” ehk merevee omadusi, milles olid juba mõnda aega olnud merisiiliku Arbacia või hulkrakne Nereis viljastamata munad, avastas Lilly, et munadest eraldub ainet, millel oli võime liimi sperma tükkideks. Täheldatud aglutinatsioon osutus liigispetsiifiliseks ja Lilly nimetas viljastamata munarakust eritatavat aglutinatsioonifaktorit viljastumisaineks või väetis(inglise keelest fertilization - fertilization). Lilly viljastamisteooria olemus seisneb äratundmises, et munaraku perifeerses piirkonnas on fertilisiini, millel on afiinsus sperma pinnaretseptorite suhtes (sperma antifertiliin). Tänu sellele afiinsusele seob väetis Lilly sõnul sperma. Kuid selleks, et pretendeerida universaalsusele ja selgitada mitte ainult sugurakkude ühinemise mehhanismi, vaid ka spermatosoidide aglutinatsiooni põhjuseid, polüspermia vältimise võimalust, viljastamisprotsessi kõrget spetsiifilisust jne, vajas väetamise teooria arvukalt eeldusi, mille alusel. mille ikke see lõpuks välja suri.
Juba varajaste viljastamise uuringute käigus tekkis idee gamonitest - ainetest, mis aktiveerivad või blokeerivad viljastamise üksikuid etappe. Vastavalt päritolule eristasid nad munade kaudu eritatavaid günogamoone ja meeste sugurakkude poolt toodetud androgamoone. Seega arvati, et munarakust difundeeruv gynogamon 1 aktiveerib seemnerakkude liikumist, ületades androgamon 1 toime, mis pärsib seemnerakkude liikumist. Gynogamon 2 on väetise sünonüüm ja androgamon 2 on sperma antifertilisiin.
20. sajandi viiekümnendatel muudeti idee väetise ja antifertilisiini koostoimest spetsiifilise fagotsütoosi hüpoteesiks. Selle kontseptsiooni kohaselt annab interakteeruvate molekulide olemasolu munaraku ja sperma pinnal täiendava tõmbluku reaktsiooni, mis võimaldab spermatosoididel munarakku imenduda.
Vaatamata teatud spekulatiivsusele mängisid need ja paljud teised sarnased hüpoteesid spermatosoidide ja munarakkude interaktsiooni mehhanismide kohta positiivset rolli, paljastades esiteks terve spetsiifiliste molekulide perekonna olemasolu interakteeruvate sugurakkude pinnal ja teiseks algatades süstemaatilise nende molekulide olemuse uurimine .
Eelmise sajandi teine pool oli ultrastruktuuri- ja molekulaarbioloogiliste uuringute hiilgeaeg, mis paljastas viljastamise ajal väga erinevaid spetsiifilisi rakkude interaktsiooni vorme. Sai selgeks, et universaalne väetamise teooria, kui see võiks eksisteerida, oleks vaid selle protsessi korraldamise kõige üldisemate põhimõtete kogum.
Konkreetsed väetamise mehhanismid sõltuvad paljudest teguritest. Piisab, kui öelda välise ja sisemise seemendusega loomade viljastamise ainulaadsuse kohta. Ilmselgelt on teatud erinevused viljastamisprotsessis tingitud ka sellest, et erinevatel loomadel toimub spermatosoidide tungimine munarakku oogeneesi erinevatel etappidel. Paljudes anneliidides, molluskites, nematoodides ja koorikloomades tungivad spermatosoidid profaasi staadiumis esimest järku munarakkudesse. Teistes anneliidides, molluskites ja putukates - primaarse munaraku metafaasi staadiumis. Paljudele selgroogsetele on iseloomulik seemendamine sekundaarse munaraku metafaasi staadiumis. Mõnedel koelenteraatidel ja merisiilikutel toimub viljastumine küpse munaraku staadiumis pärast küpsemisjaotuste lõppemist ja suuna- või redutseerimiskehade vabanemist. Lõpuks ei saa jätta meenutamata erinevaid spermatüüpe, mille hulgas on lipulisi vorme ja ilma flagellata (näiteks amööboidsed spermatosoidid), akrosoomiga ja ilma, akrosoomi niidiga ja ilma. Loomulikult erinevad igal sellisel juhul spetsiifilised mehhanismid, mis tagavad sugurakkude vahelise peene interaktsiooni.
Kui leiate vea, tõstke esile mõni tekstiosa ja klõpsake Ctrl+Enter.
Nagu teate, kogeb iga tüdruk ja seejärel iga naine pärast puberteedieas seda kord kuus. See on üsna keeruline füsioloogiline protsess, mille käigus küps munarakk vabaneb munasarjast munajuhasse. Siin toimub väetamine.
Ovulatsiooni tunnused
Sperma sulandumine munaga toimub kaheteistkümne tunni jooksul pärast selle väljumist munajuhast. Ovulatsiooni aja arvutamine pole keeruline ja üks kõige usaldusväärsemaid meetodeid selle määramiseks on temperatuur pärasooles. Seda protseduuri tuleb teha iga päev mitu kuud. Temperatuuri mõõdetakse samal ajal, varahommikul, voodist tõusmata, kasutades kõige tavalisemat termomeetrit.
Kui sisestate andmed graafikule, näete oma munaraku küpsemiskõverat. Enne menstruatsiooni algust maksimaalne temperatuur langeb ja ovulatsiooni hetk toimub kas madala temperatuuri viimasel päeval või selle tõusu esimesel päeval. Kõige soodsam päev munaraku viljastamiseks on ovulatsiooni toimumise või paar päeva enne selle algust.
Seda seletatakse asjaoluga, et munajuha õõnsusse sattunud spermatosoidid jäävad elujõuliseks mitu päeva. Teades ovulatsiooni päeva, ei saa te mitte ainult last eostada, vaid ka proovida. Selleks on olemas erinevad eostuskalendrid.
Väetamise mehhanism
Munaraku viljastamine on pikk ja keeruline mehhanism, mille käigus toimub meeste ja naiste sugurakkude ühinemine. Seemnevedelik, mis siseneb naise tuppe seksuaalvahekorra ajal, sisaldab ligikaudu 60–150 miljonit küpset spermat. Emaka pideva kokkutõmbumise tõttu haarab see aktiivselt seemnevedelikku ja seetõttu liiguvad liikuvad spermatosoidid mitmeks minutiks emakaõõnde ja jõuavad seejärel munajuha kaugematesse osadesse, kus munarakk asub.
Hoolimata asjaolust, et meeste sugurakke on palju, puutuvad nad oma teel kokku paljude takistustega (tupe happeline keskkond, emakakaela kanali limaskestade sisu jne) ning munarakku saab viljastada vaid üks kiirematest spermatosoididest. . Tõsi, arvukad uuringud on tõestanud, et munarakku võivad tungida mitmed spermatosoidid, kuid munaraku päriliku informatsiooniga tuum võib ühenduda vaid ühe sperma tuumaga, mille tulemusena moodustub vaid üks embrüo. Muidugi on juhtumeid, kus viljastamisprotsessi käigus sünnib mitu embrüot ja selle tulemusena sünnivad kaksikud.
Sperma läbib emasraku tugevaid membraane, kuna see lahustub selle pea akrosomaalses kapslis sisalduvate ensüümide toimel. Munaga kokku puutudes kapsel rebeneb ja sellest hakkab membraanidele kinnituma akrosoomniit ning eralduvad ained, mis muna membraani hävitavad. Pärast väikese ala lahustamist tungib akrosoomniit sügavale muna sisse ja ühendub tihedalt selle sisemise sisuga. Seejärel imendub sperma pea tuum ja sisemine sisu naise sugurakku.
Muutused munas
Sperma täielik tungimine naise sugurakku käivitab selle füsioloogiliste protsesside oluliste muutuste protsessi. Muna koored muutuvad palju läbilaskvamaks, mis on väga oluline toitainete aktiivseks kogumiseks, mille abil embrüo areneb. Valgud, kaltsium ja süsivesikud hakkavad aktiivsemalt tootma, kaltsiumi ja fosfori maksimaalne kogus imendub - üldiselt tehakse ettevalmistusi loote arenguks.
Kõige olulisemad ja olulisemad sündmused sündimata lapse jaoks toimuvad umbes kaheteistkümne tunni jooksul pärast sperma tungimist munarakku. Sel ajal ühinevad mees- ja naisrakkude tuumad, mis kannavad kogu pärilikku teavet. Moodustub uus rakk täiskomplektiga kromosoome, millest areneb seejärel embrüo ja lõpuks sünnib uus inimene.
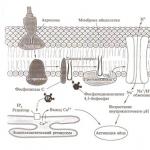
Pärast naiste suguelunditesse tungimist ilmutab sperma pärast protsessi viljastumisvõimet Kapitsatsioonid. Selle olemus: sperma peas on ensüümi glükosüültransferaasi sisaldavad alad. Kuid selle ensüümi blokeerivad galaktoos ja N-atsetüülglükoosamiin. Naiste suguelunditesse sekreteeritud glükoproteiinid vabastavad blokeerivaid ensüüme. Seejärel suudab sperma ära tunda N-atsetüülglükoosamiini jäägid pilucid tsoonis (membraan, mis on kaetud follikulaarsete rakkude kihiga). Seejärel leiab ensüüm potentsiaalse substraadi. Edasi tuleb teine protsess, mille käivitavad munarakud – Akrosomaalne reaktsioon. Selle mehhanism: pärast kokkupuudet tarretisega sisenevad Ca ioonid sperma. Välise seemenduse käigus tulevad Ca ioonid veest ja sisemise ajal endoplasmaatilisest retikulumist. Paralleelselt toimub membraaniprotsesside ümberstruktureerimise protsess, mis tagab Na ja prootonite sisenemise väljapoole. Selle tulemuseks on pH tõus, mis viib aktiini polümerisatsioonini. Järgmisena aktiveeritakse domeeni ATPaas. Seejärel toimub akrosomaalse vesiikuli eksotsütoos - topeltmembraan asendatakse ühekordsega. Valk bendiin ilmub tekkivale akrosomaalsele väljakasvule (tunneb ära munarakul olevad retseptorid). Sabaga kahepaiksetel, roomajatel ja lindudel ei tungi munasse üsna sageli mitte üks, vaid mitu spermat ning nende loomade munades on välja töötatud spetsiaalsed mehhanismid, mis inaktiveerivad liigse spermatosoidi tuumasid. Enamikul teistel selgroogsetel Polüspermia Seda takistavad pinnareaktsioonid, mis ei lase enam kui ühel spermatosoidil munarakku tungida. Selliste loomade munadel on pindmine kortikaalsete graanulite kiht; nende selgroogsete munades, mis võimaldavad mitme spermatosoidi läbitungimist, selliseid graanuleid ei ole. Selgroogsetel, kes lasevad munarakku tungida vaid ühel spermal, on esimene reaktsioon, mis tekib vastusena sperma sulandumisele munarakuga, munaraku plasmamembraani elektriliste omaduste kiire muutus. Positiivne membraanipotentsiaal takistab polüspermia teket, samas kui äsja viljastatud munaraku vähenenud potentsiaal võimaldab seda. Sperma akrosoom sisaldab hüdro- ja rotüütilisi ensüüme, näiteks akrosiini, mis sarnaneb kemotrüpsiiniga. Akrosoom sisaldab ensüümi (georuronidaasi), mis lõhustab koronaradiata. Need ensüümid tagavad spermatosoidide tungimise munarakku. Kontakttsoonis toimub munaraku ja sperma membraani lagunemine. Kontakttsoonis tekivad mitsellid tühimiku moodustumisega ja sperma sisu tungib sisse. Sündmus, mis takistab polüspermiat ja leiab aset mõni minut pärast sperma tungimist munarakku, on Kortikaalne reaktsioon. Kortikaalsed graanulid, alates kohast, kus munarakk ja sperma sulanduvad, liiguvad plasmamembraani sisepinnale, sulanduvad sellega ja vabastavad seejärel oma sisu muna ümbritsevasse ruumi. Pärast kortikaalsete graanulite sisu vabanemist blokeeritakse teiste spermatosoidide tungimine munarakku muutuste tõttu pilutsiidses tsoonis ja munaraku plasmamembraanis. Kohtumehhanism. reageerida. sarnane akrosomaalsele reaktsioonile - ensüümide eksotsütoos plasmamembraani ja vitelliini membraani vahele. Need graanulid sisaldavad polüsahhariide, mis võimaldavad vee läbitungimist. Lisaks veele sisenevad ka muud ained. Hüoliin tekitab membraanile hüoliinikihi, mis tagab purustamisel blastomeeride säilimise. Samuti moodustab see kaitse sperma eest. Imetajatel on tsoonireaktsioon, spermatosoidide sissetungimisel muutuvad munarakul olevad retseptorid ja takistavad teiste spermatosoidide tungimist. Pärast sperma tungimist munarakku geneetiline materjal dekondenseerub ja tuumamembraan hävib. Dekondenseerunud geneetilise materjali ümber moodustub uus kest. Moodustub 2 esituuma, sooritades liigutusi - protuumade tantsu. Pärast seda tuumamembraanid lagunevad ja kromosoomid dubleeritakse, millele järgneb mitootiline jagunemine. See on viimane etapp, mis takistab hübridiseerumist.
Väetamise mehhanismid
Loomade viljastamisprotsessi võib jagada kolme faasi. Esimest faasi iseloomustab spermatosoidide lähenemine munarakule enne nende kokkupuudet. Selles faasis toimuvad sugurakkude vahelised kauged interaktsioonid. Teine faas algab sperma kinnitumisega munaraku pinnale. Sel ajal täheldatakse sugurakkude vahelisi kontaktinteraktsioone. Viljastamisprotsessi kolmas faas algab pärast spermatosoidide tungimist munarakku ja lõpeb mehe ja naise sugurakkude tuumade ühinemisega. See faas iseloomustab interaktsiooni munaraku sees.
Kauged interaktsioonid sugurakkude vahel
Kauginteraktsiooni tagavad mitmed mittespetsiifilised tegurid, mille hulgas on erilisel kohal sugurakkude poolt toodetud keemilised ained. On teada, et sugurakud eritavad gamoone või sugurakkude hormoone. Munade toodetud gamoone nimetatakse günogamoonideks ja spermatosoidide toodetud gamoone nimetatakse androgamoonideks. Naiste sugurakud eristavad kahte gamoonide rühma: gynogamons I ja gynogamons II, mis mõjutavad meeste sugurakkude füsioloogiat. Spermatosoidid toodavad androgamoone I ja II.
Mõned neist kemikaalidest on loodud selleks, et suurendada tõenäosust, et sperma kohtub munaga. On teada, et spermatosoidide liikumine munarakku toimub kemotaksise kaudu - spermatosoidide liikumine mööda teatud munaraku sekreteeritavate kemikaalide kontsentratsioonigradienti. Kemotaksist on usaldusväärselt näidatud paljude loomarühmade, eriti selgrootute puhul: molluskid, okasnahksed ja poolloomad. Merisiiliku munadest on eraldatud kemotaktilised tegurid: mõnel liigil on see kümnest aminohappest koosnev peptiid, mida nimetatakse speraktiks, teistel liikidel on see neljateistkümnest aminohappest koosnev peptiid ja seda nimetatakse resactiks. Kui nende ainete ekstrakte lisatakse merevette, hakkavad vastavate liikide spermatosoidid oma kontsentratsioonigradienti ülespoole liikuma.
Imetajate spermatosoidide liikumisel mööda munajuha ülemisi osi on hädavajalik reotaksise nähtus – võime liikuda vastutulevale munajuhavedeliku voolule.
Pärast seda, kui sperma läbib munaraku kaitsvaid membraane ja puutub kokku selle plasmamembraaniga, algab sugurakkude vaheline kontaktinteraktsioon, mis viib spermatosoidide tungimiseni munaraku tsütoplasmasse.
Sugurakkude vahelised kontaktinteraktsioonid
Sperma kokkupuude munamembraaniga viib sugurakkude aktiveerumiseni. Aktiveerimisreaktsioon on seotud keeruliste morfoloogiliste, biokeemiliste ja füüsikalis-keemiliste muutustega sugurakkudes. Meeste suguraku aktiveerimine on peamiselt seotud akrosomaalse reaktsiooniga ja naissoost raku aktiveerimine kortikaalse reaktsiooniga.
Akrosoom reaktsioon mida iseloomustavad kiired muutused sperma pea akrosomaalses aparaadis, millega kaasneb selles sisalduvate spermolüsiinide vabanemine ja akrosomaalse filamendi väljutamine munaraku pinna suunas.
Vaatleme akrosomaalse reaktsiooni üldist skeemi erinevate mereselgrootute rühmade esindajatel - okasnahksed, anneliidid, kahepoolmelised, maohingajad jne.
Sperma pea ülaosas lahustub plasmamembraan ja sellega külgnev akrosoomi vesiikulite membraani osa (lüüsib). Mõlema membraani vabad servad ühinevad üheks membraaniks. Spermolüsiinid eralduvad avatud akrosoomist keskkonda ja põhjustavad munaraku membraanide lahustumist spermaga kokkupuutumise kohas. Pärast seda ulatub akrosmaalse aparaadi sisemine membraan väljapoole ja moodustab torukujulise väljakasvu (akrosoomfilament). Akrosomaalne filament pikeneb, läbib täiendavate munamembraanide lõtvunud ala ja puutub kokku muna plasmamembraaniga. Akrosomaalse hõõgniidi kokkupuutealal muna pinnaga plasmamembraanid ühinevad ja akrosomaalse toru (hõõgniidi) sisu ühendub muna tsütoplasmaga. Membraani liitmise tulemusena moodustub tsütoplasmaatiline sild. Veidi hiljem liiguvad spermatosoidi tuum ja tsentriool läbi tsütoplasma silla munaraku tsütoplasmasse. Akrosoomi reaktsioon lõpeb sperma membraani sisestamisega munamembraani. Sellest hetkest alates on sperma ja munarakk juba üks rakk (joon. 7, 8, 9.).
Joonis 7. Sperma akrosoom reaktsioon: A - B - akrosoomi välismembraani ja sperma membraani liitmine. Akrosomaalse vesiikuli sisu efusioon; 1 - akrosoommembraan; 2 - sperma membraan; 3 - globulaarne aktiin; 4 - akrosoomi ensüümid; D - E - aktiini polümerisatsioon ja akrosomaalse laienduse moodustumine; 5 - bindiin; 6 - akrosoomi kasv; 7 - aktiini mikrokiud; 8 - spermatosoidi tuum. (Golichenkovi järgi)
Vaatamata akrosomaalse reaktsiooni üldisele sarnasusele on nende loomade vahel teatud erinevused. Seega ei sisalda okasnahksete akrosoomiaparaat erinevalt ussidest ja molluskitest lüütilisi ensüüme. Enamikul uuritud loomadel moodustub üks akrosomaalne filament ja mõnel ussil mitu sellist filamenti.

Joonis 8. Akrosomaalse reaktsiooni järjestus merisiilikul.(Golichenkovi järgi)
Selgroogsetel viljastamisel toimub ka akrosoomreaktsioon. Madalamatel selgroogsetel (jänesed ja tuurad) sarnaneb see paljuski selgrootute loomade sperma aksomaalse reaktsiooniga.

Joonis 9. Muna ja sperma membraanide koosmõjul viljastamise ajal toimuvate protsesside skeem (Gilberti järgi).
Hail, roomajatel ja lindudel, kelle munad on kaetud tiheda koorega, toimub sugurakkude ühinemine enne nende kestade moodustumist. Nendel loomadel täidab akrosoom jätkuvalt oma algset rolli ja on hästi arenenud.
Imetajate akrosoomi reaktsioon erineb selgrootute ja madalamate selgroogsete omast. Imetajate spermatosoidides toimub akrosomaalne reaktsioon ilma akrosomaalse väljakasvu moodustumiseta, lähenedes munaraku pinnale, sulandub sperma oma plasmamembraaniga läbi pea külgpinna.
Putukatel ja kõrgematel kaladel toimub sugurakkude ühinemine pärast tihedate täiendavate munamembraanide täielikku moodustumist. Nendel juhtudel tungib seemnerakk munarakku mikropillikanalite kaudu ja sugurakkude ühinemine toimub ilma akrosoomi osaluseta.
Muna aktiveerimine. Kortikaalne reaktsioon. Pärast seda, kui meessoost sugurakk kinnitub munaraku pinnale ja selle akrosomaalne filament puutub kokku ooplasma pinnaga, toimub munarakk aktiveerumine. Muna aktiveerimine on seotud keerukate muutustega selle tegevuse erinevates aspektides. Aktiveerumise kõige silmatorkavam väline ilming on muutused ooplasma pinnakihis, mida nimetatakse kortikaalseks reaktsiooniks (joon. 10).

Joonis 10. Kortikaalne reaktsioon merisiiliku munas A-sperma lähendamine munarakule; B-D - kortikaalse reaktsiooni järjestikused etapid; näitab kortikaalsete graanulite sisu vabanemise lainet, mis levib sperma tungimise kohast, membraani eraldumist ja perivitelliinse ruumi moodustumist, hüaliinkihi moodustumist; gs-hüaliinkiht; Jo-kollane kest kg-kortikaalne graanul; oo-viljastusmembraan PM-plasmamembraan; pp-perivitelliini ruum, mis on täidetud perivitelliinvedelikuga (Ginzburgi järgi).
Vaatleme ajukoore reaktsiooni järjestikuseid etappe kõige põhjalikumalt uuritud merisiiliku munade näitel. Kortikaalne reaktsioon algab iga kortikaalse graanuliga piirneva membraaniga, mis kleepub munaraku plasmamembraanile. Sel hetkel graanulid avanevad ja nende sisu valatakse vitelliini membraani. Kortikaalsete graanulite sisu sekretsiooniprotsess algab seemnerakkude kinnituskohast ja levib lainetena igas suunas, kuni katab kogu munaraku pinna. Osa ajukoore graanulite sekreteeritavast sisust on hüdreeritud ja lahustunud, moodustades perivitelliinvedeliku, mis surub vitelliini membraani munaraku plasmalemmast eemale, mis viib perivitelliinse ruumi mahu suurenemiseni. Teine osa kortikaalsete graanulite sisust sulandub vitelliini membraaniga, mis pakseneb ja muundub viljastamismembraaniks. Osa kortikaalsetest graanulitest, mis ei osale viljastamismembraani moodustamises, muundatakse tihedaks kihiks, mida nimetatakse plasmamembraani kohal paiknevaks hüaliinkihiks. Kui viljastamismembraan on moodustunud, ei suuda teised spermatosoidid munaraku munaplasmasse tungida.
Viimastel aastatel on uuritud kortikaalsete graanulite sisu keemilist koostist. On tõestatud, et kortikaalsete graanulite sisu sisaldab järgmisi aineid: a) proteolüütiline ensüüm (aktelliindelaminaas), mis lõhub sidemeid rakumembraani ja munaraku plasmamembraani vahel; b) proteolüütiline ensüüm (sperma retseptori hüdrolaas), mis vabastab vitelliini membraanile ladestunud sperma; c) glükoproteiin, mis tõmbab vett vitelliini membraani ja plasmamembraani vahele, põhjustades nende eraldumise; d) väetamismembraani teket soodustav tegur; e) struktuurvalk hüaliin, mis osaleb hüaliinkihi moodustamises.
Mis on kortikaalse vastuse bioloogiline tähtsus?
Esiteks on kortikaalne reaktsioon mehhanism, mis kaitseb munarakku liigsete spermatosoidide tungimise eest.
Teiseks toimib kortikaalse reaktsiooni tulemusena tekkinud perivitelliinne vedelik spetsiifilise keskkonnana, kus toimub embrüo areng.
Muna aktiveerimisel täheldatakse muid muutusi selle tegevuse erinevates aspektides.
Esiteks väheneb meioosi blokeerinud pidur ja tuumatransformatsioonid jätkuvad just sellest etapist, kus need peatusid ajal, mil munarakk munasarjast lahkus.
Teiseks täheldatakse mitmeid biokeemilisi muutusi, millega kaasneb suurenenud süsivesikute metabolism ning suurenenud lipiidide ja valkude süntees.
Kolmandaks suureneb järsult rakumembraani läbilaskvus naatriumi- ja kaaliumiioonidele.
Sündmused, mis esinevad munas pärast sperma tungimist
Pärast seda, kui sperma akrosomaalse filamendi plasmamembraan sulandub munaraku plasmamembraaniga, kaotab sperma oma liikuvuse ja selle värbamine munarakku toimub aktiveeritud munarakust lähtuvate jõudude toimel. Tavaliselt tõmmatakse sperma munaplasmasse koos sabaga, kuid mõnikord visatakse saba ära. Kuid isegi juhtudel, kui lipp tungib muna sisse, visatakse see ära ja lahustub.
Sperma tugevalt kondenseerunud tuum hakkab paisuma, kromatiin lõdveneb ja tuum muutub omapäraseks struktuuriks, mida nimetatakse isase protuumiks.
Sarnased muutused toimuvad ka munaraku tuumas, mille tulemusena moodustub emane protuum. Protuumade moodustumise ajal toimub DNA replikatsioon mööda kromosoome. Seejärel hakkavad esituumad liikuma muna keskosa suunas. Igat esituuma ümbritsevad tuumamembraanid hävivad, protuumad lähenevad üksteisele ja tekib kariogaamia. Kariogaamia on viljastamise viimane etapp. Protuumade ühinemisel moodustub diploidse kromosoomikomplektiga tuum. Seejärel võtavad kromosoomid ekvatoriaalse positsiooni ja toimub sügoodi esimene jagunemine.
Ooplasmaatiline segregatsioon. Pärast sperma tungimist algavad munaraku tsütoplasma (ooplasma) intensiivsed liikumised. Sel juhul toimub ooplasmi erinevate komponentide eraldamine ja segunemine, mida nimetatakse ooplasmaatiliseks segregatsiooniks. Selle protsessi käigus tuuakse välja embrüo ruumilise korralduse peamised elemendid. Seetõttu nimetatakse seda arenguetappi ka promorfogeneesiks: see tähendab, et sel ajal seatakse verstapostid tulevaste morfogeneetiliste protsesside jaoks.
Mono- ja polüspermia
Ühe sperma tungimist munarakku nimetatakse füsioloogiliseks monospermiaks. Monospermia on omane kõikidele välisseemendusega loomarühmadele ja paljudele siseseemendusega loomadele (need, kellel nagu imetajatelgi on väikesed munad).
Teistel loomadel, näiteks mõnedel lülijalgsetel (putukatel), molluskitel (tigude klassi), akordaatidel (hailaadsed kalad, sabaga kahepaiksed, roomajad ja linnud) tungib munasse suur hulk seemnerakke. Seda nähtust nimetatakse füsioloogiliseks polüspermiaks. Kuid sel juhul on munaraku tuumaga ühendatud ainult ühe spermatosoidi tuum, ülejäänud aga hävivad (joon. 11).

Riis. 11. Polüspermia vesivillil. A-sperma tungimine munarakku küpsemise teise jagunemise metafaasi staadiumis; B-sünkroonsed muutused seemnetuumades, seemnetähtede teke; B-emane tuum ühendub ühe seemnetuumaga; G-E sünkarüon siseneb mitoosi, ülearvulised seemnetuumad surutakse vegetatiivsesse poolkera ja degenereeruvad. Numbrid munarakkude kujutise kohal näitavad aega pärast sperma tungimist temperatuuril 23 o (vastavalt Ginzburgile).
Füsioloogilise monospermia korral on munaraku kaitsmiseks polüspermia eest spetsiaalsed mehhanismid. Esimene mehhanism on seotud membraanipotentsiaali muutusega. On kindlaks tehtud, et konnamunas muutub mõni sekund pärast kokkupuudet spermaga membraani laeng -28-lt 8 mV-le ja püsib positiivsena 20 minutit. Samasuguseid muutusi membraanipotentsiaalis leiti ka merisiiliku munades. Selgus, et membraani positiivne laeng takistab polüspermiat. Veel üks laialt levinud mehhanism munaraku kaitsmiseks liigsete spermatosoidide tungimise eest on seotud viljastamismembraani ja perivitelliinvedeliku moodustumisega.