Citogenetski mehanizmi oplodnje. Oplodnja, njene faze, biološka suština. Kako dolazi do oplodnje jajne ćelije?
Oplodnja je složen mehanizam uzastopnih procesa, samo uz striktno poštovanje uslova koji se mogu postići željena trudnoća. Dakle, žena mora imati zrelo jaje, prohodne jajovode i dovoljnu debljinu sluzokože materice, koja mora biti spremna za pričvršćivanje oplođenog jajašca. Nakon implantacije, žensko tijelo mora osigurati hormonsku podršku za trudnoću.
Muškarac mora imati dovoljan broj morfološki normalnih pokretnih zrelih spermatozoida sposobnih da kroz cerviks prodiru u šupljinu materice, jajovod i oplode jajnu stanicu.
Kod žena se polne ćelije nazivaju jajima, a kod muškaraca spermatozoida.
Jajna stanica podsjeća na običnu ćeliju, okruglog je oblika, uključuje jezgro koje sadrži DNK (majčin genetski materijal), citoplazmu i membranu. Ćelija je okružena takozvanom „blistavom krunom“, koja se sastoji od zaštitnih ćelija.
Sperma ima fundamentalno drugačiju strukturu. To je zbog funkcije koju obavlja: potrebno je savladati udaljenost od cerviksa do jajovodnog dijela jajovoda. Spermatozoid ima glavu (koja sadrži DNK: očev genetski materijal), vrat i rep. Na glavi sperme nalazi se posebna struktura - akrosom, koji sadrži enzime koji olakšavaju prodiranje sperme u jaje.
Osnovna razlika između ženskog i muškog organizma je u tome što je opskrba jajima u ženskom tijelu strogo ograničena i polaže se tokom prenatalnog razvoja djevojčice. Kada se urođena zaliha folikula iscrpi, žena gubi reproduktivnu funkciju (menopauza). U muškom tijelu proces formiranja sperme ne ovisi o dobi i konstantan je.
Fiziološki proces oplodnje
Proces oplodnje u potpunosti ovisi o hormonskoj regulaciji, psihoemocionalnom stanju ženskog tijela, faktorima okoline, itd. Menstrualni ciklus uobičajeno počinje prvog dana menstruacije, kada je hormonska pozadina „nula“. Tada počinje rast grupe folikula, od kojih se do 5-7 dana menstrualnog ciklusa odabire jedan folikul. Kada folikul dostigne veličinu od 18-25 mm (obično 12-14 dana menstrualnog ciklusa), dolazi do ovulacije.
Kao rezultat toga, jaje se oslobađa u trbušnu šupljinu, odakle, uz pomoć fimbrija (posebnih izraslina jajovoda), ulazi u ovarijalni dio jajovoda, gdje se susreće sa spermatozoidima, koji, zauzvrat, nakon ejakulacije, putuje dug put nekoliko sati kroz ženski genitalni trakt. Kao rezultat ovog sastanka, u prisustvu povoljnih uslova sredine, dolazi do oplodnje.
Nakon toga, embrion koji se već dijeli (zigota - sadrži genetski materijal majke i oca) putuje kroz jajovode do materice zbog peristaltičkih kontrakcija zidova jajovoda, kretanja resica i kapilarnog toka tekućine. Nakon toga, ulaskom u šupljinu maternice, embrij se pričvršćuje za jedan od zidova maternice, dolazi do implantacije i daljnjeg razvoja embrija.
Neophodni uslovi za uspešan proces oplodnje:
sazrijevanje folikula u jajniku, jaje spremno za oplodnju;
ovulacija, dovoljna funkcija žutog tijela;
normalna koncentracija visokokvalitetne aktivne sperme;
prodiranje sperme u jajovode, njihovo kretanje kroz ženski genitalni trakt, sposobnost oplodnje;
Oplodnja, početni trenutak nastanka nove genetske individualnosti, je proces kombinovanja ženskih i muških polnih ćelija.
Kao rezultat oplodnje, pojavljuje se jednoćelijski embrij s diploidnim setom hromozoma i aktivira se lanac događaja koji su u osnovi razvoja organizma.
Biološki značaj oplodnje je ogroman: kao preduslov za razvoj nove individualnosti, ono je istovremeno i uslov za nastavak života i evoluciju vrste.
Treba naglasiti da oplodnja nije jednokratni čin, već proces koji traje manje-više dugo. Ovo je proces u više faza, koji razlikuje sljedeće faze: privlačenje sperme jajnom stazom, vezivanje gameta i, konačno, fuzija muških i ženskih reproduktivnih stanica. U naučnoj literaturi događaji povezani sa konvergencijom gameta ponekad se nazivaju inseminacijom, praveći razliku između vanjske i unutrašnje oplodnje, ovisno o tome da li se muške reproduktivne stanice oslobađaju u okoliš ili u ženske genitalije. Vanjska oplodnja je tipična za životinje koje žive u vodenoj sredini. Unutrašnja oplodnja karakteristična je uglavnom za kopnene životinje, iako je prilično česta među stanovnicima vodene sredine. Inseminacija može biti slobodna, pri kojoj su spermatozoidima dostupna sva područja oocita, ali može biti i ograničena kada se na površini jajne stanice nalazi gusta opna s mikropilom. Tokom unutrašnjeg osjemenjavanja kod većeg broja životinja, muške gamete se prenose na ženke u obliku spermatofore, posebne kapsule koje sadrže spermu. Spermatofori se prvo oslobađaju u okoliš, a zatim se na ovaj ili onaj način prenose u ženski reproduktivni trakt.
Veza gameta određuje mogućnost kariogamija ili nuklearna fuzija. Zahvaljujući kariogamiji dolazi do spajanja očevih i majčinih hromozoma, što dovodi do formiranja genoma nove osobe. Kao rezultat fuzije gameta, pojavljuje se diploidni zigot, obnavlja se sposobnost replikacije DNK i počinje priprema za podjele cijepanja. Mehanizmi aktivacije jajeta za razvoj su relativno autonomni. Njihovo uključivanje se može izvršiti uz oplodnju, koja se javlja, na primjer, tokom prirodnog ili umjetnog djevičanskog razvoja, ili partenogeneza.
Interesovanje za problem oplodnje daleko prevazilazi okvire same embriologije. Fuzija gameta je plodno korišten model za proučavanje finih molekularnih i ćelijskih mehanizama specifičnih interakcija ćelijskih membrana; proučavati molekularne osnove metaboličke aktivacije i proliferacije somatskih stanica. Također je od opšteg biološkog interesa da je oplodnja upečatljiv i, možda, jedinstven primjer potpunog preokreta diferencijacije ćelija. Zaista, visoko specijalizovane zametne ćelije nisu sposobne za samoreprodukciju. Oni su haploidni i ne mogu se dijeliti. Međutim, nakon fuzije oni se pretvaraju u totipotentnu ćeliju, koja služi kao izvor formiranja svih tipova ćelija svojstvenih datom organizmu.
Istorija otkrića oplodnje izgubljena je u magli vremena. U svakom slučaju, italijanski prirodnjak opat Lazzaro Spallanzani (1729-1799) je u 18. veku eksperimentalno dokazao da oplodnja zavisi od prisustva sperme i po prvi put izvršio veštačku oplodnju jaja žaba, mešajući ih sa spermom dobijenom od testise. Ipak, smisao događaja koji se dešavaju u ovom slučaju ostao je nejasan skoro sve do poslednje četvrtine 19. veka, kada je Oscar Hertwig (1849-1922) krajem 1870-ih, proučavajući oplodnju kod morskih ježeva, došao do zaključka da je suština ovaj proces je fuzija jezgara zametnih ćelija. Zajedno sa radovima Belgijanca Eduarda van Benedena (1883, okrugli crv), njemačkog naučnika Theodora Boverija (1887, okrugli crv) i švicarskog zoologa Hermanna Fohla (1887, morska zvijezda), istraživanje O. Hertwiga postavilo je temelje modernim idejama o đubrenje. Treba naglasiti da su upravo ovi radovi poslužili kao čvrsta osnova za pretpostavku da je nukleus nosilac nasljednih svojstava. Upravo je T. Boveri (1862-1915), u nizu briljantnih citoloških studija, potkrijepio teoriju individualnosti hromozoma kasnih 1880-ih i stvorio osnovu citogenetike.
Ubrzo nakon što je suština oplodnje razjašnjena, istraživači su svoju pažnju usmjerili na mehanizme koji leže u osnovi ovog procesa. Ova oblast istraživanja ostaje aktuelna i danas. Vodstvo u razvoju teorije oplodnje pripada američkom istraživaču Franku Lillyju (1862-1915). Proučavajući svojstva “vode za jaja”, odnosno morske vode u kojoj su neko vrijeme bila prisutna neoplođena jaja ježa Arbacia ili poliheta Nereis, Lilly je otkrila da se iz jajašca oslobađa supstanca koja ima sposobnost lepiti spermu u grudvice. Pokazalo se da je uočena aglutinacija specifična za vrstu, a Lilly je faktor aglutinacije koji luči neoplođeno jaje nazvala oplodnjom, ili đubrivo(od engleskog fertilization - oplodnja). Suština Lillyjeve teorije oplodnje je u spoznaji da se u perifernom dijelu jajne stanice nalazi fertilizin, koji ima afinitet za površinske receptore sperme (sperma antifertilizin). Zahvaljujući ovom afinitetu, fertilizin vezuje, prema Lili, spermu. Međutim, da bi se tvrdila univerzalnost i objasnila ne samo mehanizam spajanja gameta, već i razlozi aglutinacije spermatozoida, mogućnost prevencije polispermije, visoka specifičnost procesa oplodnje itd., teoriji oplodnje su bile potrebne brojne pretpostavke, pod jaram od kojeg je na kraju izumrla.
Već u toku ranih studija oplodnje pojavila se ideja o gamonima - tvarima koje osiguravaju aktivaciju ili blokiranje pojedinih faza oplodnje. U skladu sa porijeklom, razlikovali su ginogamone koje luče jajašca i androgamone koje proizvode muške reproduktivne stanice. Stoga se vjerovalo da gynogamon 1, difundirajući iz jajne stanice, aktivira kretanje spermatozoida, prevladavajući djelovanje androgamona 1, koji inhibira kretanje spermatozoida. Gynogamon 2 je sinonim za fertilizin, a androgamon 2 je antifertilizin sperme.
Pedesetih godina 20. stoljeća ideja o interakciji fertilizina sa antifertilizinom transformirana je u hipotezu specifične fagocitoze. Prema ovom konceptu, prisustvo molekula u interakciji na površini jajne ćelije i sperme obezbeđuje komplementarnu reakciju zatvarača koja omogućava da se sperma apsorbuje u jaje.
Uprkos određenoj spekulativnosti, ove i mnoge druge slične hipoteze o mehanizmima interakcije između spermatozoida i jajne ćelije odigrale su pozitivnu ulogu, otkrivajući, prvo, postojanje čitave porodice specifičnih molekula na površini interakcionih gameta i, drugo, inicirajući sistematsko istraživanje prirode ovih molekula.
Druga polovina prošlog veka bila je vrhunac ultrastrukturnih i molekularno-bioloških istraživanja, koja su otkrila širok spektar specifičnih oblika ćelijskih interakcija tokom oplodnje. Postalo je jasno da bi univerzalna teorija oplodnje, da postoji, bila samo kao skup nekih od najopštijih principa za organizaciju ovog procesa.
Specifični mehanizmi oplodnje zavise od mnogih faktora. Dovoljno je reći o jedinstvenosti oplodnje kod životinja s vanjskom i unutarnjom oplodnjom. Očigledno je da su određene razlike u procesu oplodnje također posljedica činjenice da se kod različitih životinja prodiranje sperme u jajnu ćeliju događa u različitim fazama oogeneze. Kod mnogih anelida, mekušaca, nematoda i rakova, sperma prodire u oocite prvog reda u fazi profaze. Kod ostalih anelida, mekušaca i insekata - u fazi metafaze primarne oocite. Mnoge kralježnjake karakterizira osjemenjivanje u fazi metafaze sekundarne oocite. Kod nekih koelenterata i morskih ježeva do oplodnje dolazi u fazi zrelog jajeta nakon završetka podjela sazrijevanja i oslobađanja usmjerenih ili redukcijskih tijela. Konačno, ne može se ne prisjetiti raznolikosti vrsta spermatozoida, među kojima postoje flagelarni oblici i spermatozoidi bez flagela (na primjer, ameboidne spermatozoide), sa i bez akrozoma, sa i bez akrozomalne niti. Naravno, u svakom takvom slučaju se razlikuju specifični mehanizmi koji osiguravaju suptilnu interakciju između zametnih stanica.
Ako pronađete grešku, označite dio teksta i kliknite Ctrl+Enter.
Kao što znate, nakon puberteta, svaka djevojčica, a zatim i svaka žena, to doživi jednom mjesečno. Ovo je prilično složen fiziološki proces tokom kojeg se zrela jajna stanica oslobađa iz jajnika u jajovod. Ovdje dolazi do oplodnje.
Karakteristike ovulacije
Fuzija spermatozoida sa jajetom se dešava u roku od dvanaest sati nakon njegovog izlaska iz jajovoda. Nije teško izračunati vrijeme ovulacije, a jedna od najpouzdanijih metoda za određivanje je, odnosno temperatura u rektumu. Ovaj postupak se mora provoditi svakodnevno nekoliko mjeseci. Temperatura se mjeri u isto vrijeme, rano ujutro, bez ustajanja u krevetu, najobičnijim termometrom.
Ako unesete podatke u grafikon, možete vidjeti krivulju sazrijevanja vašeg jajeta. Prije početka menstruacije, maksimalna temperatura se smanjuje, a trenutak ovulacije nastupa ili posljednjeg dana niske temperature ili prvog dana njenog porasta. Najpovoljniji dan za oplodnju jajne ćelije je onaj kada nastupi ovulacija, ili nekoliko dana pre njenog početka.
To se objašnjava činjenicom da spermatozoidi koji su ušli u šupljinu jajovoda ostaju održivi nekoliko dana. Znajući dan ovulacije, možete ne samo začeti dijete, već i pokušati. U tu svrhu postoje različiti kalendari začeća.
Mehanizam oplodnje
Oplodnja jajne ćelije je dug i složen mehanizam tokom kojeg dolazi do spajanja muških i ženskih reproduktivnih ćelija. Semenska tečnost, koja ulazi u žensku vaginu tokom seksualnog odnosa, sadrži otprilike 60 do 150 miliona zrelih spermatozoida. Zbog kontinuirane kontrakcije maternice, sjemena tekućina se njome aktivno hvata, pa se pokretni spermatozoidi nekoliko minuta kreću u šupljinu maternice, a zatim dospiju do udaljenih dijelova jajovoda, gdje se nalazi jajna stanica.
Unatoč činjenici da postoji mnogo muških reproduktivnih stanica, one na svom putu nailaze na mnoge prepreke (kisela sredina vagine, mukozni sadržaj cervikalnog kanala i tako dalje), a samo jedan od najbržih spermatozoida može oploditi jajnu stanicu. . Istina, brojne studije su dokazale da nekoliko spermatozoida može prodrijeti u jajnu stanicu, ali jezgro s nasljednom informacijom jajne stanice može se povezati s jezgrom samo jednog spermatozoida, što rezultira formiranjem samo jednog embrija. Naravno, postoje slučajevi kada se tokom procesa oplodnje proizvede nekoliko embriona, a kao rezultat se rađaju blizanci.
Spermatozoid prolazi kroz jake membrane ženske ćelije zbog rastvaranja enzima sadržanim u akrozomalnoj kapsuli njene glave. Dolazeći u kontakt s jajetom, kapsula puca, a iz nje se akrosomalna nit počinje vezivati za membrane i oslobađaju se tvari koje uništavaju membranu jajeta. Otapajući malo područje, akrosomalna nit prodire duboko u jaje i čvrsto se povezuje s njegovim unutarnjim sadržajem. Zatim se jezgro i unutrašnji sadržaj glave sperme apsorbiraju u žensku reproduktivnu ćeliju.
Promjene u jajetu
Potpuni prodor sperme u žensku reproduktivnu ćeliju pokreće proces značajnih promjena u fiziološkim procesima u njoj. Ljuske jajeta postaju znatno propusnije, što je vrlo važno za aktivno nakupljanje hranjivih tvari uz pomoć kojih će se embrij razvijati. Proteini, kalcij i ugljikohidrati počinju se aktivnije proizvoditi, apsorbira se maksimalna količina kalcija i fosfora - općenito se izrađuju pripreme za razvoj fetusa.
Najvažniji i najznačajniji događaji za nerođeno dijete dešavaju se u roku od oko dvanaest sati nakon što sperma prodre u jajnu stanicu. U ovom trenutku se ujedinjuju jezgre muških i ženskih ćelija koje nose sve nasljedne informacije. Formira se nova ćelija sa punim setom hromozoma, iz kojih će se potom razviti embrion i na kraju će se roditi nova osoba.
Nakon prodiranja u ženski genitalni trakt, sperma pokazuje sposobnost oplodnje nakon procesa Kapocitacije. Njegova suština: glava sperme ima područja koja sadrže enzim glikoziltransferazu. Ali ovaj enzim blokiraju galaktoza i N-acetilglukozamin. Glikoproteini koji se izlučuju u ženski genitalni trakt oslobađaju enzime za blokiranje. Tada sperma može prepoznati ostatke N-acetilglukozamina u pilucidnoj zoni (membrana prekrivena slojem folikularnih stanica). Enzim tada pronalazi potencijalni supstrat. Slijedi drugi proces, iniciran membranama jajeta - Akrosomalni reakcija. Njegov mehanizam: nakon kontakta sa želeom, Ca joni ulaze u spermu. Prilikom eksterne inseminacije Ca joni dolaze iz vode, a kod unutrašnje oplodnje iz endoplazmatskog retikuluma. Paralelno se odvija proces restrukturiranja membranskih procesa koji osiguravaju ulazak Na i protona u van. To dovodi do povećanja pH, što dovodi do polimerizacije aktina. Zatim se aktivira domen ATPase. Tada dolazi do egzocitoze akrozomalne vezikule - dvostruka membrana se zamjenjuje jednom. Protein bendin se pojavljuje na nastaloj akrosomalnoj izraslini (prepoznaje receptore na jajetu). Kod repatih vodozemaca, gmizavaca i ptica ne jedan, već nekoliko spermatozoida često prodire u jaje, a jajašca ovih životinja razvila su posebne mehanizme koji inaktiviraju jezgre viška sperme. Kod većine drugih kralježnjaka Polispermija Sprečavaju ga površinske reakcije koje sprečavaju više od jednog spermatozoida da prodre u jajnu stanicu. Jaja takvih životinja imaju površinski sloj kortikalnih granula; u jajima onih kičmenjaka koji omogućavaju prodiranje nekoliko spermatozoida, takvih granula nema. Kod kičmenjaka, koji dozvoljavaju samo jednom spermiju da prodre u jajnu stanicu, prva reakcija koja se javlja kao odgovor na fuziju spermatozoida sa jajetom je brza promjena električnih svojstava plazma membrane jajeta. Pozitivan membranski potencijal sprečava nastanak polispermije, dok smanjen potencijal u novooplođenoj jajnoj stanici to omogućava. Akrosom sperme sadrži hidro- i rotelitičke enzime, na primjer, akrozin, sličan hemotripsinu. Akrosom sadrži enzim (georuronidazu) koji razdvaja korona radijatu. Ovi enzimi osiguravaju prodiranje sperme u jaje. U kontaktnoj zoni dolazi do raspadanja membrane jajne ćelije i sperme. U kontaktnoj zoni formiraju se micele sa stvaranjem jaza i sadržaj sperme prodire unutra. Događaj koji sprečava polispermiju i javlja se nekoliko minuta nakon što sperma prodre u jajnu stanicu je Kortikalna reakcija. Kortikalne granule, počevši od tačke u kojoj su se jajna ćelija i spermatozoid spojili, kreću se do unutrašnje površine plazma membrane, spajaju se sa njom, a zatim ispuštaju svoj sadržaj u prostor koji okružuje jaje. Nakon oslobađanja sadržaja kortikalnih granula, prodiranje drugih spermatozoida u jaje blokirano je promjenama u pilucidnoj zoni i plazma membrani jajeta. Sudski mehanizam. reagovati. slična akrosomalnoj reakciji - egzocitoza enzima u prostor između plazma membrane i vitelne membrane. Ove granule sadrže polisaharide koji omogućavaju prodiranje vode. Osim vode, ulaze i druge tvari. Hyolin stvara hiolinski sloj na membrani, koji osigurava zadržavanje blastomera tokom drobljenja. Takođe stvara zaštitu od sperme. Sisavci imaju zonsku reakciju, kada spermatozoidi prodru unutra, receptori na jajnoj stanici se menjaju i sprečavaju prodiranje drugih spermatozoida. Nakon što sperma prodre u jajnu stanicu, genetski materijal se dekondenzira i nuklearna membrana se uništava. Oko dekondenzovanog genetskog materijala formira se nova ljuska. Formiraju se 2 pronukleusa, izvodeći pokrete - ples pronukleusa. Nakon toga, nuklearne membrane se raspadaju i hromozomi se dupliciraju, nakon čega slijedi mitotička podjela. Ovo je posljednja faza koja ima prepreku za hibridizaciju.
Mehanizmi oplodnje
Proces oplodnje kod životinja može se podijeliti u tri faze. Prvu fazu karakterizira približavanje sperme jajnoj stanici prije njihovog kontakta. Tokom ove faze odvijaju se udaljene interakcije između zametnih ćelija. Druga faza počinje pričvršćivanjem sperme na površinu jajne ćelije. U ovom trenutku se uočavaju kontaktne interakcije između zametnih ćelija. Treća faza procesa oplodnje počinje nakon što spermatozoid prodre u jajnu stanicu i završava spajanjem jezgara muške i ženske zametne stanice. Ova faza karakteriše interakciju unutar jajeta.
Udaljene interakcije između zametnih ćelija
Udaljene interakcije osiguravaju se brojnim nespecifičnim faktorima, među kojima posebno mjesto zauzimaju hemijske supstance koje proizvode zametne ćelije. Poznato je da reproduktivne ćelije luče gamone ili hormone gameta. Gamoni koje proizvode jajašca nazivaju se ginogamoni, a oni koje proizvode spermatozoidi nazivaju se androgamoni. Ženske zametne ćelije razlikuju dvije grupe gamona: ginogamone I i ginogamone II, koje utječu na fiziologiju muških zametnih stanica. Spermatozoidi proizvode androgamone I i II.
Neke od ovih hemikalija su dizajnirane da povećaju verovatnoću da se spermatozoidi sretnu sa jajetom. Poznato je da se kretanje spermatozoida do jajne ćelije odvija putem kemotaksije - kretanja spermatozoida duž gradijenta koncentracije određenih hemikalija koje izlučuje jajna ćelija. Hemotaksa je pouzdano dokazana kod mnogih grupa životinja, posebno beskičmenjaka: mekušaca, bodljokožaca i poluhordata. Iz jaja morskih ježeva izdvojeni su hemotaktički faktori: kod nekih vrsta to je peptid koji se sastoji od deset aminokiselina i naziva se speract, kod drugih vrsta je peptid koji se sastoji od četrnaest aminokiselina i naziva se resact. Kada se ekstrakti ovih supstanci dodaju u morsku vodu, spermatozoidi odgovarajuće vrste počinju da se kreću gore u gradijentu koncentracije.
U kretanju spermatozoida sisara duž gornjih delova jajovoda, bitan je fenomen reotakse - sposobnost kretanja protiv nadolazećeg toka tečnosti jajovoda.
Nakon što sperma prođe kroz zaštitne membrane jajne ćelije i dođe u kontakt sa njenom plazma membranom, počinju kontaktne interakcije između zametnih ćelija koje će dovesti do prodiranja sperme u citoplazmu jajne ćelije.
Kontaktne interakcije između zametnih ćelija
Kontakt spermatozoida sa membranom jajne ćelije dovodi do aktivacije zametnih ćelija. Reakcija aktivacije povezana je sa složenim morfološkim, biohemijskim i fizičko-hemijskim promjenama u zametnim stanicama. Aktivacija muške zametne stanice prvenstveno je povezana s akrosomalnom reakcijom, a ženske s kortikalnom reakcijom.
Akrosomska reakcija karakterizira brze promjene u akrosomalnom aparatu glave spermatozoida, praćene oslobađanjem spermolizina sadržanih u njemu i izbacivanjem akrozomalnog filamenta prema površini jajeta.
Razmotrimo opću shemu akrosomalne reakcije kod predstavnika različitih grupa morskih beskralježnjaka - bodljokožaca, anelida, školjkaša, gastro-dihača itd.
Na vrhu glave sperme, plazma membrana i susjedni dio membrane akrozomalnih vezikula se rastvaraju (liziraju). Slobodni rubovi obje membrane spajaju se u jednu membranu. Spermolizini se oslobađaju iz izloženog akrozoma u okolinu i dovode do rastvaranja jajnih membrana na mjestu kontakta sa spermom. Nakon toga, unutrašnja membrana akrozmalnog aparata strši prema van i formira izraslinu u obliku cijevi (akrosomalni filament). Akrosomalni filament se izdužuje, prolazi kroz olabavljeno područje dodatnih membrana jajeta i dolazi u kontakt sa plazma membranom jajeta. U području kontakta akrosomalnog filamenta s površinom jajeta, plazma membrane se spajaju i sadržaj akrosomalne cijevi (filamenta) povezuje se s citoplazmom jajeta. Kao rezultat fuzije membrane, formira se citoplazmatski most. Nešto kasnije, jezgro i centriol sperme proći će kroz citoplazmatski most u citoplazmu jajne stanice. Akrozomska reakcija završava umetanjem membrane spermatozoida u membranu jajeta. Od ovog trenutka, spermatozoid i jajna ćelija su već jedna ćelija (sl. 7, 8, 9.).
Fig.7. Akrosomska reakcija spermatozoida: A - B - fuzija vanjske membrane akrozoma i membrane spermatozoida. Izlijevanje sadržaja akrozomalne vezikule; 1 - akrozomska membrana; 2 - membrana sperme; 3 - globularni aktin; 4 - akrozomski enzimi; D - E - polimerizacija aktina i formiranje akrozomalnog nastavka; 5 - bindin; 6 - rast akrozoma; 7 - aktinski mikrofilamenti; 8 - jezgro sperme. (prema Golichenkovu)
Unatoč općoj sličnosti akrosomalne reakcije, postoje određene razlike među njima kod ovih životinja. Dakle, kod bodljokožaca, za razliku od crva i mekušaca, akrosomalni aparat ne sadrži litičke enzime. Kod većine proučavanih životinja formira se jedan akrozomalni filament, a kod nekih crva i nekoliko takvih filamenata.

Fig.8. Redoslijed akrosomalne reakcije kod morskog ježa.(prema Golichenkovu)
Tokom oplodnje kod kičmenjaka dolazi i do akrozomske reakcije. Kod nižih kralježnjaka (lamreys i jesetra) u mnogočemu je slična aksomalnoj reakciji sperme beskičmenjaka.

Fig.9. Šema procesa koji se javljaju tokom interakcije membrana jajne ćelije i sperme tokom oplodnje (prema Gilbertu).
Kod ajkula, gmizavaca i ptica, čija su jaja prekrivena gustom ljuskom, spajanje gameta se događa prije nego što se ove ljuske formiraju. Kod ovih životinja akrosom nastavlja da ispunjava svoju prvobitnu ulogu i dobro je razvijen.
Akrozomski odgovor kod sisara razlikuje se od beskičmenjaka i nižih kralježnjaka. Kod spermatozoida sisara akrosomalna reakcija se odvija bez formiranja akrozomalnog izraslina.Približavajući se površini jajeta, spermatozoid se spaja sa svojom plazma membranom kroz bočnu površinu glave.
Kod insekata i viših riba, spajanje zametnih stanica događa se nakon što se potpuno formiraju guste dodatne membrane jaja. U tim slučajevima, spermatozoid prodire u jajnu stanicu kroz mikropilarne kanale i spajanje gameta se dešava bez učešća akrozoma.
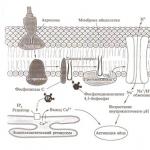
Aktivacija jaja. Kortikalna reakcija. Nakon što se muška reproduktivna stanica veže za površinu jajeta i njen akrozomalni filament dođe u kontakt s površinom ooplazme, dolazi do aktivacije jajeta. Aktivacija jajeta povezana je sa složenim promjenama u različitim aspektima njegove aktivnosti. Najupečatljivija vanjska manifestacija aktivacije su promjene u površinskom sloju ooplazme, koje se nazivaju kortikalna reakcija (slika 10).

Slika 10. Kortikalni odgovor u jajetu morskog ježa A-približavanje sperme jajetu; B-D - uzastopne faze kortikalne reakcije; prikazuje val oslobađanja sadržaja kortikalnih granula, širenje od mjesta prodiranja sperme, odvajanje membrane i formiranje perivitelinskog prostora, formiranje hijalinskog sloja; gs-hijalinski sloj; Jo-žumance ljuske kg-kortikalne granule; oo-oplodna membrana PM-plazma membrana; pp-perivitelinski prostor ispunjen perivitelinskom tečnošću (prema Ginzburgu).
Razmotrimo uzastopne faze kortikalne reakcije na primjeru najpotpunije proučavanih jaja morskog ježa. Kortikalna reakcija počinje tako što se membrana koja graniči sa svakom kortikalnom granulom prianja na plazma membranu jajeta. U ovom trenutku se granule otvaraju i njihov sadržaj se izlije u vitelnu membranu. Proces lučenja sadržaja kortikalnih granula počinje od mjesta pričvršćivanja sperme i širi se valovito u svim smjerovima dok ne pokrije cijelu površinu jajne stanice. Dio izlučenog sadržaja kortikalnih granula se hidratizira i otapa, formirajući perivitelinsku tekućinu, koja gura vitelinsku membranu od plazmaleme jajeta, što dovodi do povećanja volumena perivitelinskog prostora. Drugi dio sadržaja kortikalnih granula spaja se sa vitelnom membranom, koja se zgušnjava i pretvara u oplonu za oplodnju. Neke od kortikalnih granula koje ne učestvuju u formiranju membrane za oplodnju transformišu se u gust sloj koji se naziva hijalinski sloj koji se nalazi iznad plazma membrane. Jednom kada se formira membrana za oplodnju, drugi spermatozoidi ne mogu prodrijeti u ooplazmu jajne stanice.
Poslednjih godina proučavan je hemijski sastav sadržaja kortikalnih granula. Pokazalo se da sadržaj kortikalnih granula sadrži sledeće supstance: a) proteolitički enzim (aktelin delaminaza), koji razbija veze između ćelijske membrane i plazma membrane jajeta; b) proteolitički enzim (hidrolaza receptora sperme), koji oslobađa spermu taloženu na vitelnoj membrani; c) glikoprotein koji uvlači vodu u prostor između vitelne membrane i plazma membrane, uzrokujući njihovo razdvajanje; d) faktor koji podstiče formiranje opne za oplodnju; e) strukturni protein hijalin, uključen u formiranje hijalinskog sloja.
Koji je biološki značaj kortikalnog odgovora?
Prvo, kortikalna reakcija je mehanizam koji štiti jajnu stanicu od prodora prekobrojnih spermatozoida.
Drugo, perivitelinska tekućina nastala kao rezultat kortikalne reakcije služi kao specifično okruženje u kojem se odvija razvoj embrija.
Kada se jaje aktivira, uočavaju se druge promjene u različitim aspektima njegove aktivnosti.
Prvo, kočnica koja je blokirala mejozu se smanjuje i nuklearne transformacije se nastavljaju od same faze u kojoj su prestale u trenutku kada je jaje napustilo jajnik.
Drugo, uočava se niz biohemijskih promjena, praćenih pojačanim metabolizmom ugljikohidrata i povećanom sintezom lipida i proteina.
Treće, propusnost stanične membrane za jone natrijuma i kalija naglo se povećava.
Događaji koji se javljaju u jajetu nakon prodiranja spermatozoida
Nakon što se plazma membrana akrozomalnog filamenta spermatozoida spoji sa plazma membranom jajne ćelije, spermatozoid gubi svoju pokretljivost i dolazi do njegovog regrutacije u jajnu ćeliju usled dejstva sila koje izlaze iz aktiviranog jajeta. Obično se sperma uvlači u ooplazmu zajedno sa repom, ali ponekad se rep odbacuje. Međutim, čak i u slučajevima kada flagelum prodre u jaje, on se odbacuje i rastvara.
Visoko zgusnuto jezgro sperme počinje da bubri, hromatin se popušta i jezgro se pretvara u osebujnu strukturu koja se zove muški pronukleus.
Slične promjene se javljaju u jezgri jajeta, što rezultira formiranjem ženskog pronukleusa. Tokom formiranja pronukleusa dolazi do replikacije DNK duž hromozoma. Nakon toga, pronukleusi počinju da se kreću prema centru jajeta. Nuklearne membrane koje okružuju svaki od pronukleusa su uništene, pronukleusi se zbližavaju i dolazi do kariogamije. Kariogamija je posljednja faza oplodnje. Kada se pronukleusi spoje, formira se jezgra sa diploidnim skupom hromozoma. Tada kromosomi zauzimaju ekvatorijalni položaj i dolazi do prve podjele zigote.
Ooplazmatska segregacija. Nakon prodiranja sperme, počinju intenzivni pokreti citoplazme jajne stanice (ooplazma). U ovom slučaju dolazi do razdvajanja i miješanja različitih komponenti ooplazme, što se naziva ooplazmatska segregacija. Tokom ovog procesa ocrtavaju se glavni elementi prostorne organizacije embriona. Stoga se ova faza razvoja naziva i promorfogenezom: to znači da se u ovom trenutku postavljaju prekretnice za buduće morfogenetske procese.
Mono- i polispermija
Prodor jednog spermatozoida u jajnu stanicu naziva se fiziološka monospermija. Monospermija je karakteristična za sve grupe životinja sa spoljašnjom oplodnjom i mnoge životinje sa unutrašnjom oplodnjom (one koje, kao i sisari, imaju mala jaja).
Kod drugih životinja, na primjer, kod nekih člankonožaca (insekata), mekušaca (klasa puževaca), hordata (ribe slične ajkuli, repati vodozemci, gmizavci i ptice), veliki broj spermatozoida prodire u jaje. Ovaj fenomen se naziva fiziološka polispermija. Međutim, u ovom slučaju samo jezgro jednog spermatozoida je povezano sa jezgrom jajne ćelije, dok su ostali uništeni (Sl. 11).

Rice. 11. Polispermija u tritonu. A-prodiranje sperme u jaje u fazi metafaze druge podjele sazrijevanja; B-sinhrone promjene u jezgri sjemena, formiranje sjemenih zvijezda; B-žensko jezgro se povezuje s jednim od sjemenih jezgara; G-E sinkarion ulazi u mitozu, prekobrojna sjemena jezgra se potiskuju u vegetativnu hemisferu i degeneriraju. Brojevi iznad slike jajnih ćelija označavaju vrijeme nakon prodiranja spermatozoida na temperaturi od 23 o (prema Ginzburgu).
Kod fiziološke monospermije postoje posebni mehanizmi zaštite jajne stanice od polispermije. Prvi mehanizam je povezan s promjenom membranskog potencijala. Utvrđeno je da se u jajetu žabe, nekoliko sekundi nakon kontakta sa spermom, naboj membrane mijenja sa -28 na 8 mV i ostaje pozitivan 20 minuta. Iste promjene u membranskom potencijalu pronađene su i u jajima morskog ježa. Pokazalo se da pozitivan naboj membrane sprečava polispermiju. Još jedan široko rasprostranjen mehanizam za zaštitu jajne ćelije od prodiranja prekobrojnih spermatozoida povezan je sa formiranjem opložne membrane i perivitelinske tečnosti.