Mis on mootori üksus. Mootori üksuste klassifikatsioon. Ristresistentne südamelihas
Peamine morpho funktsionaalne element neuromuskulaarse aparaadi skeleti lihaste on mootoriüksus (DE). See sisaldab seljaaju foleeroni oma Axon innervrored lihaste kiud. Lihaste sees moodustab see akson mitu otsa oksi. Iga selline oksi moodustab kontakt - neuro-lihaste sünapsi eraldi lihaskiud. Mettoyeroni tulevad närviimpulssid põhjustavad teatud lihaskiudude rühma vähenemise. Väikeste lihaste mootoriühikud, kes kasutavad õhukeseid liikumisi (silmalihased, harjad), sisaldavad väikest kogust lihaskiudude. Suurema sadade korda rohkem. Kõik DE sõltuvalt funktsionaalsetest omadustest jaguneb kolmeks rühmaks:
I. Aeglane väsimatu. Neid moodustavad "punased" lihaskiud, kus vähem kui müofibrillid. Nende kiudude vähenemise määr ja tugevus on suhteliselt väikesed, kuid nad on vähe väsinud. Seetõttu nimetatakse nad toonikuks. Selliste kiudude lühendite reguleerimine viiakse läbi väikese arv modeons, kelle aksonitel on vähe otsakambreid. Näide - Cambalo-sarnane lihas.
Ib Kiire, kergesti väsinud. Lihas kiud sisaldavad palju müofibrilli ja neid nimetatakse "valgeks". Kiiresti lõigata ja arendada suuremat tugevust, kuid kiiresti rehve. Seetõttu nimetatakse neid faasi. Nende de suurimate failide modeons on paljude lõppvarude rasva akson. Nad tekitavad suure sageduse närviimpulsse. Lihase silmad.
IIa. Kiire, vastupidav väsimusele. Hõivata vahepealne asend.
Sile lihaste füsioloogia
Sujuvad lihased on saadaval kõige seedetrakti elundite, laevade, erinevate näärmete, kuseteede väljundkanalite seintes. Nad on tahtmatud ja tagavad seedehäirete ja kuseteede ja kuseteede personteeritud, säilitades laevade tooni. Erinevalt skeleti, silelihaseid moodustavad rakkude sagedamini kui spindli kujulised ja väikesed suurused, millel ei ole põiki eraldisi. Viimane on tingitud asjaolust, et tellija ei ole tellitud struktuuri. Myofibrillis koosneb aktiini peenematest tüvedest, mis lähevad erinevates suundades ja kinnitades erinevate saitide Sarchatimmas. Üksi protofibrillid asuvad Aktinovi kõrval. Sarkoplasmaatilise reticulumi elemendid ei moodusta torude süsteemi. Eraldi lihasrakud kombineeritakse madalate elektriliste resistentsete kontaktide madalate elektriliste resistentsusega, mis tagab erutuse jaotuse kogu sujuva lihaste struktuuri vältel. Eraldatavus ja juhtivus silelihaseid on madalamad kui skeletil.
Membraani potentsiaal on 40-60 mV, kuna MMC membraanil on naatriumioonide suhteliselt suur läbilaskvus. Lisaks paljud siledad lihased MP ei ole konstantne. See väheneb perioodiliselt ja naaseb uuesti algtaseme juurde. Sellised võnkumised nimetatakse aeglaseks laineteks (MV). Kui Vertexi aeglased lained jõuavad depolarisatsiooni kriitilisele tasemele, hakkavad sellega kaasnema lühenditega kaasnevad potentsiaalid (joonis). MV ja PD hoitakse sujuvatel lihastel kiirusel ainult 5 kuni 50 cm / s. Selliseid silelihaseid nimetatakse spontaanselt aktiivseks, st Neil on automatiseerimine. Näiteks sellise tegevuse tõttu esineb soole peristalistlik. Instituutide rütmi draiverid asuvad vastavate soolede esialgses osakondades.
PD põlvkond MMC-s on tingitud nende kaltsiumi ioonide sisendist. Elektromehaanilised konjugeerimismehhanismid on samuti erinevad. Vähendamine areneb rakus sisalduva kaltsiumi kulul PD-ga, vahendab kaltsiumi ühendamist, millel on lühendavamad miofibrillid kõige olulisem rakkude valk - Calmodulin.
Vähendamise kõver on ka erinev. Varjatud periood, lühendamisperiood ja eriti lõõgastumine on palju kauem kui skeletilihastes. Vähendamine kestab paar sekundit. Sujuvad lihased, erinevalt skeleti, plasttooni omapärane nähtus. See võime on pikka aega vähendamisseisundis ilma märkimisväärse energiatarbimise ja väsimuseta. Tänu sellele varale säilitatakse siseorganite vorm ja laevade toon. Lisaks on silelihasrakud ise tõmberetseptorid. Kui nad on pingeid, hakkavad PD loodud, mis toob kaasa MMC vähenemise. Seda nähtust nimetatakse müogeense mehhanismi kontraktiivse tegevuse reguleerimiseks.
Liikumine on vajalik tingimus organismi arendamiseks ja olemasoluks, selle juurdepääs keskkonnale. See on liikumine, mis on aluseks sihipärase käitumise, mis ilmutab sõnad Na Kornettein: "ilmselge tohutu bioloogilise tähtsusega seotud organismide on peaaegu ainult vorm rakendamise mitte ainult suhtlemise keskkonnaga, Kuid ka aktiivne mõju sellele keskkonnale, mis muudab seda üksikisikutele mitteseotumatu. Tulemused ... ". Teine liikumiste olulisuse ilming on see, et mis tahes erialase tegevuse keskmes on lihaste töö.
Kõik mootori sordid viiakse läbi kasutades lihas-. See moodustab spetsialiseerunud anatoomiliste üksuste: lihased, skelett ja kesknärvisüsteem.
Teatava kaaluga lihvimihaldussüsteemis eraldatakse passiivne osa - skelett ja lihaste aktiivne osa.
Skelett sisaldab luud ja nende ühendused. (Näiteks liigesed).
Skelett See toimib toetuse siseorganeid, lihaste kinnitamise koht kaitseb siseorganeid väliste mehaaniliste kahjustuste eest. Luu-luuüdi asub luude luud - vere moodustuva keha. Lugudes on suur hulk mineraalaineid (kõige esitatud kaltsiumi, naatriumi, magneesiumi, fosfor, kloori). Luu on dünaamiline reaalajas koe, millel on kõrge tundlikkus erinevate regulatiivsete mehhanismide suhtes endo- ja eksogeensete mõjude suhtes. Luu ei ole mitte ainult tugikeha, vaid ka mineraalvahetuse kõige olulisem osaleja (rohkem - metabolismiosas). Luude metaboolse aktiivsuse lahutamatu näitaja on luukonstruktsioonide aktiivse ümberkorraldamise ja uuendamise protsessid. Need protsessid on ühelt poolt oluline mehhanism mineraalse homeostaasi säilitamiseks, teisest küljest pakuvad luu struktuurne kohandamine muutuvatele töötingimustele, mis on eriti oluliselt tingitud tavapärasest füüsilisest kultuurist ja spordist. Boneerroika pidevate protsesside keskmes seisneb luurakkude aktiivsus - osteoblastide ja osteoklastide aktiivsus.
Lihased Vähenemise võime tõttu juhtivad keha eraldi osad liikuma ja tagama ka kindlaksmääratud asendi säilitamise. Lihaste vähendamine on kaasas suure hulga soojuse väljatöötamine ja seetõttu on soojuse põlvkonna kaasatud töölihased. Hästi arenenud lihased on suurepärased siseorganite, laevade ja närvide kaitse.
Luud ja lihased, nii massina kui ka mahu poolest moodustavad olulise osa kogu organismis, on nende suhe olulisi seksuaalseid erinevusi. Täiskasvanud mehe lihaseline kaal - 35-50% (sõltuvalt lihaste arendamisest) kogu kehakaalust on naistel umbes 32-36%. Sportlastes, mis on spetsialiseerunud tugevuse spordile, võivad lihasmassi ulatuda 50-55% -ni ja kulturistides - 60-70% kehakaalu kogumassist. Luud moodustasid 18% kehakaalust meestel ja naistel 16%.
Isik eristab kolme tüüpi lihaseid:
põiki skeleti lihaste;
põikige südamelihase;
Sujuvad lihased Siseorganid, nahk, laevad.
Sujuvad lihasedjagunevad toonik(Ei suuda arendada "kiiresti" kärpeid õõnsate organite sfinkrites) ja phazno Tonic (mis on jagatud omavamaks automatiseerima. Võime spontaanse põlvkonna faasi vähendamise. Näide võib olla seedetrakti ja ureterjalite lihased ja mitte omada - arterite lihaste kiht, seemnekanalid, silmade iirise lihased, vähendavad autonoomse närvisüsteemi impulsside mõjul. Sile lihaste mootorsõidukite innervatsioon viiakse läbi vegetatiivse närvisüsteemi rakkude protsessi abil, tundlik - seljaaju gangli rakkude protsess. Reeglina ei saa sujuva lihase vähendamist põhjustada meelevaldselt, selle lühendite reguleerimisel ei osale aju koor. Sujuvate lihaste funktsioon on pika pinge säilitamine, samas kui nad kulutavad 5-10 korda vähem ATP-d, mis vajavad sama probleemi skeletilihaste puhul.
Sujuvad lihased pakuvad õõnsate organite funktsiooni, nende seinad moodustavad. Tänu sujuvatele lihastele eksiili sisu Põie, sisikondade, kõhuga, sapipõie, emaka. Sujuvad lihased pakuvad sfinother funktsioon - Loo tingimused teatud sisu salvestamiseks põrandal (uriin põis, lootele emakas). Verelaevade kliirensi muutmine, sujuvad lihased kohandavad piirkondlikku verevoolu hapniku- ja toitainete kohalikele vajadustele, osaleda hingamise reguleerimisel bronhivaugu lumeni muutuse tõttu.
Skeleti lihased Nad on aktiivse osa lihaskonna lihaskonna süsteemi, pakkudes sihipäraseid tegevusi, peamiselt tõttu meelevaldseid liikumisi (rohkem üksikasju nende struktuuri ja tööpõhimõtete peetakse allpool).
Lihaste kiudude tüübid
Lihased koosnevad erineva tugevuse, kiiruse ja vähendamise kestusega lihastest, samuti väsimusest. Neis ensüümidel on erineva aktiivsusega ja on esitatud erinevates isomeersetes vormides. See on märgatav hingamisteede ensüümide sisalduse eristamiseks - glükoliitse ja oksüdatiivse sisaldus. Müofibrillide suhe, mitokondrid ja müoglobiin eristavad nn nn valge, punane ja vahepealsed kiud . Funktsionaalsete omaduste kohaselt jagatakse lihaskiud kiire, aeglane ja vahepealne . Kui lihaste kiud erinevad atraktsiooni aktiivsusest, varieerub hingamisteede ensüümi aktiivsuse aste üsna oluliselt, mistõttu on olemas koos valge ja punane, vahepealsed kiud.
Kõige selgemalt lihaskiud erinevad müosiini molekulaarse organisatsiooni eripärades. Tema erinevate isovormide hulgas on kaks peamist - "Fast" ja "aeglane". Histokeemiliste reaktsioonide seadmisel eristuvad nad atphustaaktiivsusega. Need omadused korreleerub hingamisteede ensüümide aktiivsusega. Tavaliselt B. kiired kiud (FF kiud - kiiresti lõigatakiire tõmblema kiud Sisse aeglane kiudLibendatud kiudude (ST) kiudude (aeglane tõmbekiud), vastupidi, oksüdatiivsete ensüümide aktiivsuse kõrgemal, on nad rikkamad müoglobiini, vaadake rohkem punast. Need on kaasatud koormustega 20-25% piires maksimaalsest tugevusest ja eristuvad hea vastupidavusega.
FT - kiud, mis võrreldi punaste kiududega väikese müoglobiini sisuga, iseloomustab kõrge kontraktiilne kiirus ja võime arendada suuremat võimsust. Võrreldes aeglase kiududega võivad nad olla kaks korda kiiremini ja arendada 10 korda suuremat jõudu. FT kiud, omakorda jagunevad FTO ja FTG kiududeks. Olulised erinevused loetletud lihasskiudude tüüpide vahel määratakse energia tootmise meetod (joonis 2.1).
Joonis fig. 2.1 Energiavarustuse erinevused eri liiki lihaskiududes (aadressil http://medi.ru/doc/g740203.htm).
Energia saamine FTO kiududes esineb samal viisil nagu st-kiud, peamiselt oksüdatiivse fosforüülimisega. Tulenevalt asjaolust, et see lagunemisprotsess jätkub suhteliselt majanduslikult (iga glükoosi molekuli puhul, koguneb 39 energia fosfaatühendi energia tootmiseks energia tootmiseks), on FTO FIBERS-il suhteliselt kõrge väsitatavuse resistentsus. Energia akumulatsioon FTG-kiududes esineb peamiselt glükolüüsi abil, s.o glükoos hapniku puudumisel puruneb suhteliselt rikas laktaadi energiat. Tänu asjaolule, et see lagunemisprotsess on ebamajanduslik (ainult 3 glükoosi molekuli jaoks kogunevad energiafosfaatühendid), on FTG kiud suhteliselt kiiresti väsinud, kuid siiski on neil võimalik arendada suurt jõudu ja reeglina sisse lülitada Submaximal ja maksimaalsed lihaste kokkutõmbed.
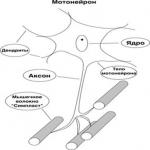
Mootoriüksused
Skeletilihaste närvilise lihasparaadi peamine morfofunktsionaalne element on lihasüksus - DE.(Jn2.2.).

Joonis 2.2. Lihasüksus
DE Sisaldab seljaaju Moteroni oma Axon innervrates lihaskiududega. Lihaste sees moodustab see akson mitu otsa oksi. Iga selline oksi moodustab kontakt - neuro-lihaste sünapsi eraldi lihaskiud. Mettoyeroni tulevad närviimpulssid põhjustavad teatud lihaskiudude rühma vähenemise. DE väikesed lihased, kes kasutavad õhukeseid liikumisi (silmalihased, harjad), sisaldavad väikest kogust lihaskiudude. Suurtes lihastes neist sadu kordi rohkem.
De aktiveeri vastavalt seadusele "kõik või mitte midagi.". Seega, kui seljaaju saadeti seljaaju eesmise hornereoni kehast impulsi närviliinidel, siis ta reageerib või kõik lihaste kiud de, Või mitte üks. Bitseps puhul tähendab see järgmist: närvisüsteemi impulss kõik kontraktiilsed elemendid (müofibrillid) kõik (ligikaudu 1500) lihaskiudude vastavad vajaliku tugevuse lühendada.
Kõik DE sõltuvalt funktsionaalsetest omadustest jaguneb kolmeks rühmaks:
I. Aeglane viga. Neid moodustavad "punased" lihaskiud, kus vähem kui müofibrillid. Nende kiudude vähenemise määr ja tugevus on suhteliselt väikesed, kuid nad on vähe väsinud, nii et need kiud kuuluvad toonikuni. Selliste lühendite reguleerimine, kiud viiakse läbi väikese mootorrataste arvuga, mille aksonitel on väikesed terminalid oksad. Näide - Cambalo-sarnane lihas.
II V. Kiire, kergesti väsinud. Lihas kiud sisaldavad palju müofibrilli ja neid nimetatakse "valgeks". Kiiresti lõigata ja arendada suuremat tugevust, kuid kiiresti rehve. Seetõttu kutsutakse neid faas. Nende de suurimate failide modeons on paljude lõppvarude rasva akson. Nad tekitavad suure sageduse närviimpulsse. Näiteks silma lihased.
II A. Kiire, vastupidav väsimusele (vahepealne).
Kõik lihaskiud ühe de viitavad sama tüüpi kiudude (FT- või ST-Fiber).
Väga täpsete ja diferentseeritud liikumiste toimivusega seotud lihased (näiteks silmade või sõrmede lihased) on tavaliselt valmistatud suurest kogusest de (1500 kuni 3000). Sellisel del on väike kogus lihaskiudude (8-50). Lihased, kes täidavad suhteliselt vähem täpseid liikumisi (näiteks jäsemete suured lihased), on oluliselt väiksem arv de, kuid nende koostis sisaldab suurt hulka kihti (600-2000).
Keskmiselt on inimesel umbes 40% aeglane ja 60% kiiretest kiududest. Kuid see on keskmine väärtus (üle skeleti lihaste), teevad lihased ka erinevaid funktsioone. Lihaste kvantitatiivne ja kvaliteetne koostis on heterogeenne, nende hulka kuuluvad mitmesugused mootoriüksused, mille suhe on ka erinev ( lihaste kompositsioon). Sellega seoses on ebavõrdse erineva lihaste kontraktsioonivõimelised võimed. Silmade välisagelad, mis pööravad silmamuna pöörama maksimaalset pinget ühe vähendamise ühe vähendamisega ainult 7,5 ms kestus, on juhusliku osa anti-gravitatsioonipõhimõte, arendab väga aeglaselt maksimaalset pinget 100 ms jaoks. Lihased, kes täidavad suuremat staatilist tööd (cambalo-like lihaste), on sageli suur hulk aeglase stiki kiude ja peamiselt dünaamiliste liikumiste tegemise lihaseid (biitseps) on suur hulk FT kiude.
Lihaste kiudude peamised omadused (sellest tulenevalt ja nende mootorite üksused), mis on määratletud ka filmide omadustega, esitatakse tabelis 1.
Vähendamise suurus (lihasjõud) sõltub lihaste morfoloogilistest omadustest ja füsioloogilisest seisundist:
1. lihaste esialgne pikkus (puhata pikad). Lihaste vähendamise võimsus sõltub lihaste algsest pikkusest või puhkepikkust. Mida tugevam lihaste venitatud üksi, seda tugevam on lühend (Frank-Starling seadus).
2. lihaste läbimõõdu või ristlõige. Rasked kaks diameetrit:
a) Anatoomiline läbimõõt - lihaste ristlõige.
b) Füsioloogiline läbimõõt - iga lihaskiudude risti ristlõige. Mida suurem füsioloogiline osa, suurem jõud omab lihaseid.
Lihasjõud mõõdetakse kõrguse või maksimaalse pinge tõstetud maksimaalse koormuse massist, mida ta suudab arendada isomeetrilises vähendamisel. Mõõdetakse kilogrammides või Newtones. Lihaskoogede mõõtmise meetod nimetatakse dünamomeetriaks.
Tõsised kahte tüüpi lihasjõudu:
1. Absoluutne jõud - maksimaalse tugevuse suhe füsioloogilisele läbimõõdule.
2. Suhteline jõud - maksimaalse tugevuse suhe anatoomilisele läbimõõdule.
Lihaste lõikamisel on see võimeline tööd tegema. Lihaste töö mõõdetakse tõstetud lasti toode lühendamise suuruse järgi.
Lihaste tööd iseloomustab võimsus. Lihasvõimsus määratakse töö suurusega seadme kohta ja mõõdetakse vattides.
Suurim töö ja võim saavutatakse keskmise koormusega.
Motooneurone koos grupiga lihaste kiudude neid on mootoriüksus. Axon Motiononov saab filiaali ja innerveerida lihaskiudude rühma. Niisiis võib üks axon innerveerida 10-3000 lihaskiududest.
Eristage mootori üksused struktuuris ja funktsioonides.
Struktuuris jagatakse mootori üksused:
1. Väikesed mootoriüksused, millel on väikemootor ja õhukese aksoni, mis on võimelised 10-12 lihaskiudude innerveerima. Näiteks nägu lihased, sõrmelihased.
2. Suure mootori üksusi esindavad suured liiderooni asutused, paks akson, mis on võimeline rohkem kui 1000 lihaskiudude innerveerima. Näiteks neljas lihases.
Funktsionaalse väärtuse abil jagatakse mootoriüksused:
1. Aeglane mootoriüksused. Nende hulka kuuluvad väikesed mootoriüksused, on kergesti häirivad, mida iseloomustab madal ergastamise kiirus, töö kõigepealt, kuid nad on praktiliselt väsinud.
2. Kiire mootori üksused. Need koosnevad suurest mootoriüksustest, halvasti põnevusest, on suur ergastamise kiirus. Neil on kõrge tugevus ja reageerimise kiirus. Näiteks poksijalihased.
Need mootori üksuste omadused on tingitud mitmest omadustest.
Lihaste kiud, mis on kaasatud mootorüksustes on sarnased omadused ja erinevused. Niisiis on aeglane lihaste kiud:
1. Rich kapillaarvõrgustik.
4. Need sisaldavad paljusid rasvu.
Tänu nendele eripäradele on nendel lihaste kiududel suur vastupidavus, mis on võimelised väikeste lühenditega, vaid kaua aega.
Kiire lihaskiudude eristusvõime:
2. omama suuremat kiirust ja vähendamise jõudu.
Seoses nende omadustega on kiire lihaskiud kiiresti väsinud, kuid neil on palju jõudu ja kõrget reageerimismäära.
Morfofunktsionaalsete omaduste kohaselt jagatakse mootori üksused 3 tüüpi:
1. aeglane rahulik de. Moldightsil on madalaim aktiveerimislävi, mis suudab säilitada kümnete minutite (st väsimatute) kümnete minutite heitmete stabiilse sageduse. Axonsil on väike paksus, väikese ergastamise kiirus, innerveerida väikese rühma lihaskiudude. Lihaste kiud arendavad väike jõudu, vähendades samal ajal nende väiksemate kontraktsioonete arvu kohalolekut - müofibrillid. Need on nn "punased kiud" (värv on tingitud kapillaarvõrgust ja väikesest hulgast müofibrillidest). Nende kiudude vähenemise määr on 1,5 - 2 korda väiksem kui kiire. Nad on väsimatult hästi arenenud kapillaarvõrgustiku tõttu suure hulga mitokondrite ja oksüdatiivsete ensüümide kõrge aktiivsusega.
2. Kiire, kergesti väsinud de. Neil on suurim kiirtee, millel on kõrgeim erutuse künnis, ei ole võimelised säilitama pikka aega stabiilset heitmete sagedust (väsinud). Paksud aksonid, millel on närvimpulsside suur kiirus, innerveerib palju lihaseid. Lihas kiud sisaldavad suurt hulka müofibrillite, nii et kui lõikamine areneb suurema võimsusega. Kõrge ensüümi aktiivsuse tõttu on vähendussagedus kõrge. Need kiud on kiiresti väsinud, sest Need sisaldavad vähem, võrreldes aeglase, mitokondritega ja neid ümbritsevad vähem kapillaarid.
3. Kiire, vastupidav väsimusele. Tugevad kiired lõikamisviidid, millel on suur vastupidavus aeroobsete ja anaeroobsete energiatootmisprotsesside kasutamise võimaluse tõttu. FIBERS 2 ja 3 tüüpi nimetatakse "valged kiud" tõttu suur sisaldus müofibrillide ja madala mooglobiini.
Aeglaste ja kiirete lihaste kiudude võrdlus
Isiku skeleti lihaste koosneb kolmest kiududest, kuid nende suhe võib sõltuvalt lihasfunktsioonist oluliselt erineda, samuti kaasasündinud ja omandatud individuaalsus. Mida rohkem valge kiudude lihaseid, seda parem inimene on kohandatud töö tegemiseks, mis nõuab kiiret kiirust ja tugevust. Punane kiudude ülekaalus annab pikaajalise töö eest vastupidavust.
Skeletilihaste struktuur
Karskeli lihas koosneb mitmesugustest lihaste kiududest, mis asuvad ühises ühendatud konteineeritud caks ja on kinnitatud skeletiga seotud kõõluste suhtes. Iga lihaskiud on õhuke (10 kuni 100 μM), mis on venitatud pikkus (5 kuni 400 mm) multi-südamiku haridus - sülastama.
Lihaskiudmembraanid on närvisüsteemi struktuuris sarnased, kuid see annab regulaarselt T-kujuline vahutamine. Lihaste kiudude sees paralleelselt membraaniga on hargnenud suletud toru süsteem - sarkoplasmaatiline reticulum - intratsellulaarne depot Ca 2+. T-System ja CP külgnevad IT-ergastamise ülekandeseadmega lihaskiudmembraaniga lõikamisstruktuurid (miofibrillos). Lihaskiudude sarkoplasmas näete põiki vahelduvaid valgus- ja tumedaid alasid vastavalt J-(isotroopsetele) ja a- (anisotroopiliste) plaate. Naabruses Myofibrillides asuvad samad kettad samal tasemel, mis annab kiudaineid ristmenetlus.Kompleks ühest tumedast ja kahest idanemisest selle kõrval, kerged kettad, piiratud põiki Z-kirjed, nimetatakse saarmur.
Iga müofibrill koosneb nende paljudest paralleelidest paks (mosic) ja Õhuke (Aktinov) Valgu niidid - miofilaments. Kiudude paksude ja õhukeste lõnga ristlõike osas asuvad kuuskantvõrgu sõlmedes kõrgelt organiseeritud. Iga paks keerme ümbritseb kuus õhuke, iga õhuke niit sõltub osaliselt keskkonda kolme naabruses paksuse. Müosiini niidid on nende ristsuunad nende peadega, mis koosnevad umbes 150 myozini molekulist. Actine niit koosneb kahest keeratud ühe ümber teise ahelate (nagu keerdunud helmed niidid) aktiin molekulid. Molekulid on paigutatud aktiini niididele troponin, Ja kahe niidi vaheliste soonte aktiini vahel tropomüoosina.
Muscle Fiber Reduction mehhanism
1954. aastal leidis G. Khaksli ja N. Khenson, et Actiin ja müosiini filamendid ei muuda oma pikkust sarjuturi lühendamisel või pikendamisel ja välja toonud slip-niitide teooria: Lihaste vähendamine toimub järjestikuse seondumise mitmekeskuse seondumise seondumise seondumise üksi ristsilda teatud piirkondades aktiini filamentide.
Ülejäänud lihaste kiududes asuvad tropomüoosi molekulid üksi selliselt, et seda takistatakse müosiini põiksuunas sildade kinnitamisega acini niidile (lihaste lõdvestunud).
Saadud ACS-Somaatilised Somaatilised PD-d jaotatakse läbi T-Tube süsteemi sügavale kiududele, põhjustades sarkoplasmaatiliste reticulumi mahutite depolariseerimist (CA2 + depoo). Membraani aktiveerimisel avaneb CP CA-kanalid ja CA2 + saagis kontsentratsiooni gradiendi juures.
Kui CA2 + ioonide kontsentratsioon kasvab, ühendab see troponiiniga, viimane vastab ja liigutab tropomüoosi niidi, avades ühendamise võimaluse üksi pea. Pea ühend põhjustab silla terava "paindumise" ja aktiini hõõgniidi liikumise 1 etappi (20 nM või 1% aktiini pikkusest), millele järgneb lõhe sild.
RE-ergastamise puudumisel CA2 + kontsentratsioon SA-pumba tilkade töö tõttu. Seetõttu on CA2 + troponinist ja tropomüasiinist uuesti blokeeritud aktiini. Samal ajal kulutatakse 1 ATP molekuli energia ühe silla ühele tööle liikumisele, veel üks - tagastada 2 Ca 2+ ioonid mahutitele.
Kõik see toob kaasa lihaste lõõgastumiseni kuni närviimpulsside järgmise voolu hetkeni, kui ülalkirjeldatud protsess korratakse.
Kombinatsioon protsesside, mis määravad jaotus PD sügavale lihaskiududesse, saagise väljund Ca2 + ioonide Sarkoplasmaatilisest reticulum, suhtlemist kontraktsiooni valkude ja lühendamine lihaskiud on kutsutud elektromehaaniline konjugatsioon.
Lihasmehaanika. Füüsikalised omadused ja lihaste kokkutõmbed
Skeleti lihaste füüsikalised omadused
1. Venivus- lihaste võime muuta selle pikkust selle tugevuse tegevuse all.
2. Elastsus - lihaste võime võtta oma esialgse pikkuse pärast tõmbe- või deformeeruva jõu lõpetamist.
3. Jõud Lihased. Seda määrab maksimaalne last, mida lihas suudab tõsta. Erijõud on maksimaalne lasti, mida lihas suudab tõsta, jagada selle füsioloogilise ristlõike ruudu sentimeetrite arvu.
4. Lihasvõime tööd tegema. Lihaste töö määrab tõstetud kauba toode lifti kõrgusele. Lihaste töö suureneb järk-järgult lasti suurendamisega, vaid teatud piirini, mille järel toob kaasa lasti suurenemine töö vähenemiseni, kuna lasti tõstmise kõrgus väheneb. Järelikult tehakse lihaste maksimaalne töö keskmiste koormuste juures ( keskmise koormuse seadus).
Lihaste kärped
Isotoonilised, isomeetrilised ja segatud lihaste kontraktsioonirežiimid eristatakse.
Jaoks isotooniline Lihaste vähendamine toimub selle pikkuse muutmine ja pinge jääb konstantseks. Selline vähendamine toimub juhul, kui lihas ei liigu lasti. Looduslikes tingimustes on keele lihaste lühendid isotoonilise lühendite tüüpi lähedal.
Jaoks isomeetrilinelihaskiudude pikkuse vähendamine jääb konstantseks, lihaste pingete muutused. Selliseid lihaste vähenemist saab talumata lasti tõsta.
Üldiselt organismi lõikamise lihaseid ei ole kunagi puhtalt isotooniline või isomeetriline, nad on alati segatud, see tähendab, muutumine ja pikkus ja lihaspinge tekib. Seda vähendusrežiimi nimetatakse auxotooniline Kui lihaspinge valitseb või aukomeetriline Kui lühendab lühendamist.
|
Kiire |
Aeglane |
|
Neuron |
|
|
Suured motneloonid |
Väikesed motneloonid |
|
Põnevus vähem |
Eruseisund rohkem |
|
Axoni läbimõõt on rohkem |
Axoni läbimõõt on väiksem |
|
Ergastamise kiirusvaade |
Ergastamise määr Vähem |
|
Sagedus suurem |
Sagedus vähem |
|
Lihaskiud |
|
|
Aktiivsus Actomosic ATPaas |
Tegevustooliline atPaas Vähem |
|
Ülaltoodud assomootiliste kiudude pakendamise tihedus |
Tektomoseliste kiudude pakendi tihedus on väiksem |
|
Saccoplasmaatiline reticulum (kaltsiumi depoo) on rohkem väljendunud |
Vähem ekspresseeritud sarkoplasmaatiline reticulum (kaltsiumi depoo) |
|
Varjatud periood pärast pd vähem |
Varjatud periood pärast PD vastuvõtmist rohkem |
|
Kaltsiumipumba tihedus |
Kaltsiumipumba tihedus Vähem |
|
Kiiremini vähendatud ja lõdvestunud |
Aeglasem vähenemine ja lõdvestunud |
|
Glükolüüsiensüümide aktiivsuse kohal |
Oksüdeerimisnsüümide aktiivsuse kohal |
|
Kiirem ATF taastumine |
ATF taastamise aeglasem, kuid ökonoomsem |
|
1 mol glükoos -2-3 moolid ATP |
1 mol glükoos 36-58 mooli ATP |
|
Moodustatakse värvimata substraadid, "kattuvad" - kiire väsimus |
väsimus on vähem väljendunud |
|
Kapillaaride suur tihedus on hapnikku, rohkem Mioglobiini |
|
|
Lihasüksus |
|
|
Vähem meelelahutuslik suur tugevus ja vähendamise määr, suur väsimus, madal vastupidavus |
Rohkem segatud, väiksema võimsusega, vähendamise määra, madal väsimus, kõrge vastupidavus |
|
sprinter | |
|
Puusade välistingimustes, aeglane kiud 13 kuni 96% -ni |
Kolmepuhastus 33% õlalihase, topeltpeaga 49%, esiküljele 46%, cambalo-kujuline 84% |
Elektromograafia meetodi neurofüsioloogilised alused.
Elektromiograafia - see meetod neuromuskulaarse süsteemi uurimiseks, registreerides lihaste elektrilised potentsiaalid. Kuigi esmakordselt elektromogrammi (EMG) registreeriti abiga telefoniseadme ne Vvensensky tagasi 1884. aastal ja 1907. aastal oli võimalik rakendada inimese EMG graafilist sisenemist, elektromüograafika intensiivset arengut kliinilise diagnostilise tehnikana algas 30-40-e 20. sajandi aastate aastate jooksul, mis on selle valdkonna teatud edusammude viivitus võrreldes näiteks elektroencefalograafia arenguga, seletavad suured nõudmised registreerimise kvaliteedi ja täpsusega tegelike parameetrite reprodutseerimise kvaliteedi ja täpsusega Elektrilised potentsiaalid elektromüograafias. Kvaliteetsete võimendite loomine, mis annavad kõrge sageduse vahemikus lineaarseid omadusi ja katoodiregistreerimismeetodite väljatöötamist, mis tagavad elektriliste potentsiaali kõrgsageduslike komponentide reprodutseerimiseta reprodutseerimisele vahemikku 20 000 Hz-le, viinud kliinilise kasutamise valdkonnas olulistele edusammudele. elektromüograafika
Intratsellulaarse registreerimisega näeb tegevuspotentsiaal välja nagu positiivne tipp, mis koosneb kiirest depolarisatsioonist, mis kestab umbes 1 ms, kiire repolarisatsiooni, mis kujutab endast potentsiaali tagastamist peaaegu ülejäänud tasemeni, kestnud umbes 2 ms; Seejärel järgitakse aeglase repolarisatsiooni, väikese jälgi hüperpolarisatsiooni ja tagastamise potentsiaali puhkuse tasemele. Kliinilises elektromüograafias, kusjuures rakuvälise registreerimisega makroelectro on esindatud lihaste kiudude potentsiaali negatiivse piik 1-3 ms.
Juht- ja registreerimise tehnika EMG
Põhimõtted Tehnika juhtiv ja registreerimine EMG ei erine seadmete elektroencefalograafia, elektrokardiograafia ja muude elektrooniliste meetodite. Süsteem koosneb elektroodidest, mis eemaldavad lihaste potentsiaalid, nende potentsiaalide võimendi või salvestusseadme. Elektromiograafia kasutab kahte tüüpi elektroode - pinna ja nõela. Pinna elektroodid on metallplaadid või kettad umbes 0,2 - 1 cm2, tavaliselt paigaldatud paarikaupa kinnitamispadjadesse, pakkudes eralduvate elektroodide vahemaad, mis on oluline registreeritud tegevuse amplituudi hindamiseks. Sellised elektroodid on lihasmootoripunkti pindala peal nahale ületatud. Nahk pühitakse alkoholiga enne elektroodi ülekatte ja seda niisutatakse naatriumkloriidi isotoonilise lahusega. Elektroodi kinnitatakse lihaste kohal, kasutades kummist triibude, manseti või leheloplastika. Kui teil on vaja naha ja elektroodi kontakti pindala pikkust uuringut, rakendatakse elektroektograafiat spetsiaalset elektroodipasta. Pinna elektroodi lihaste suur suurus ja kaugus võimaldab teil registreeruda ainult lihaste kogu aktiivsusega, mis on paljude sadu ja isegi tuhandete lihaste kiudude toimepotentsiaali sekkumine. Suure tugevdamise ja tugevate lihaste kontraktsioonide puhul registreerib pinnaelektrood naaberlihaste aktiivsust. Kõik see ei võimalda uurida individuaalsete lihaste potentsiaali parameetreid pinna elektroode abil. Saadud vastuvõtmisel hinnatakse ligikaudu ligikaudu EMG sagedust, sagedust ja amplituudi. Pinnaelektroodide eeliseks on atraumaatiline, nakkuse oht, elektroodide lihtsus. Uuringu valutu ei kehtestata mingeid piiranguid uuritud uuritavate lihaste arvu piiranguid, muudab selle meetodi laste, samuti spordi meditsiinis või massiivse ja tugeva liikumise abil seotud füsioloogilise kontrolli all oleva füsioloogilise kontrolli all.
Nõela elektroodid on kontsentrilised, bipolaarsed ja monopolaarsed. Esimeses teostuses on elektroodi esindatud õõnsa nõelaga, mille läbimõõt on umbes 0,5 mm läbimõõduga, mille sees on see eraldatud traat eraldatud sellest eraldatud plaatina või roostevabast terasest kiht. Võimalikku erinevust mõõdetakse nõela korpuse ja keskvarrast otsa vahel. Mõnikord, et suurendada paikkonna tera, nõel on isoleeritud ka väljaspool ja ainult selle elliptiline pind jäetakse nõudma viilutasandil. Standardse kontsentrilise elektroodi aksiaalse varraadi tühjenduspinna pindala on 0,07 mm 2, EMG-potentsiaalide parameetrid viitavad selle tüübi ja suuruse elektroodele. Mis märkimisväärne kasv kontaktpiirkonnas tühjenemise elektroodi, potentsiaalsed parameetrid võivad oluliselt erineda. Sama viitab muutustega elektroodi konstruktsiooni (bipolaarne monopolaarne, mitme elektroodi). Bipolaarne elektrood sisaldab kahest nõela sama isoleeritud varrast üksteisest, alasti nõuandete vahel, mis mõõdetakse millimeetri kümnendikku, mõõdetakse võimaliku erinevusega. Lõpuks kasutatakse monolaari administratsioone, elektroode kasutatakse, mis on nõel, mis on isoleeritud kogu, välja arvatud terava otsa, tühjaks 1-2 mm. Nõela elektroode kasutatakse pd individuaalsete de ja lihaskiudude parameetrite uurimiseks. Nõela elektroodi väljalaskmine on peamine esmase lihaste ja neuromuskulaarsete haiguste kliinilise missiooni orienteeritud diagnoosimisel. Üksikute PDS-i salvestamine de ja lihaste kiududes võimaldab teil täpselt hinnata kestust, amplituudit, kuju ja faasi potentsiaali
Ülesandeliigid
Sõltumata elektroodide liigist eristatakse kaks juhtiva elektrimeetodi meetodit - mono ja bipolaarne. Elektromiograafias nimetatakse monopolaari selliseks juhtimiseks, kui üks elektrood asub otseste lihaste uuritava ala lähedal ja teine \u200b\u200b- selles piirkonnas eemaldatud piirkonnas (nahk üle luu, kõrva jne. ). Monopolaarse juhtimise eeliseks on võime määrata uuringu struktuuri potentsiaali kujul ja võimaliku kõrvalekaldumise tegelik faas. Puuduseks on see, et suur kaugus elektroodide vahel, teiste lihaste osakondade potentsiaalid või isegi teistest lihastest sekkub. Bipolaarne heakskiidu on selline plii, milles mõlemad elektroodid on tihedalt lähedased ja sama kauguse uurimise all olevast lihast. Selline on juhtiv bipolaarsete või kontsentriliste nõela elektroodide abil ja ühe plokis salvestatud pinna elektroodidega. Bipolar toob kaasa madalad aste registrite aktiivsus kaugete potentsiaalide allikatest, eriti nõela elektroodide kasutamisel. Mõju mõlema elektroodi allikast pärinevate tegevuste potentsiaali erinevusele põhjustab potentsiaali vormi moonutamist ja potentsiaali tegeliku faasi kindlaksmääramist. Sellegipoolest muudab kõrge paikkonna tase see meetod kliinilises praktikas eelistas. Kuna pinna elektroodide väljalaskmine igal juhul registreerib paljude omavahel seotud PD DE häirete aktiivsus, ei ole sellise monopolaarse plii kasutamine mõtet.
Lisaks elektroodidele on võimalik EMG-võimendi sisendile kaasas potentsiaalne erinevus, maapinna pinnaelektrood on paigaldatud uuringu nahale, mis on kinnitatud elektroomi elektroodi paneeli sobiva terminali külge. Elektroodide võimalikku erinevust toidetakse pingepliiandi sisendisse. Võimendi on varustatud astmelise kasumi lülitiga, mis võimaldab reguleerida amplifikatsioonitaset sõltuvalt salvestatud aktiivsuse amplituristist. Tugevdatud elektriline aktiivsus eemaldatakse mitte ainult ostsilloskoopis, vaid ka valjuhääldis, mis võimaldab hinnata elektrivõimalusi kuulmiseks
EMG ja elektromüograafilise semiootika analüüsimise üldpõhimõtted.
Elektromüograafilise kõvera analüüs hõlmab lihaste tegelike elektriliste potentsiaali diferentseerumist võimalikest artefaktidest esimeses etapis ja seejärel peamises etapis EMG hindamise ise. Esialgne operatiivhinnang viiakse läbi ostsilloskoobi ekraanil ja akustiliste nähtuste kohta, mis tulenevad tugevdatud EMG toodangust valjuhääldile; Lõplik analüüs EMG kvantitatiivse iseloomuga ja kliinilise järeldusega tehakse paberi või filmi kirjutamise teel.
ERMG-i artefaktide potentsiaali nimetatakse potentsiaalideks, mis ei ole seotud lihaste elementide aktiivsusega. Pealiskaudse loomise korral võib esemeid olla põhjustatud elektroodi liikumisest selle lahtise lukustuse tõttu nahale, mis toob kaasa suure amplituudide väljanägemise vale kuju potentsiaali hüppab. Kõige nõepuude loovutamisega võivad elektroodi puudutamisel tekkida sarnased võimalikud muudatused, mis puudutavad juhtmete ühendavaid juhtmeid, massiivseid lihaste liikumist uuringus. Kõige levinum takistus on 50 Hz filmimine tööstusliku tööstuse seadmetest. See on kergesti tunnustatud iseloomuliku sinusoidse vormi ja konstantse sageduse ja amplituudi abil. See võib olla seotud suure elektroodiresistentsusega, mis nõuab nõelaktroodi sobivat töötlemist. Pinna elektroodidega saab põrandakate kõrvaldamist saavutada põhjalikuma naha puhastamisega alkoholiga, kasutades elektroodipasta.
EMG-analüüs sisaldab hinnangut kujul, amplituudi ja kestuse potentsiaali individuaalsete lihaste kiudude ja de ja omaduste häirete tegevuse tulenevad meelevaldse lihaseisu vähendamise. Lihaspotentsiaali eraldi kõikumise vorm võib olla mono-, di- Trio või polüfhaas. Nagu elektroencefalograafias, nimetatakse monofaasilist sellist võnkumist, kus kõver teeb kõrvalekalde ühes suunas isoelektrilisest joonest ja naaseb esialgse tasemeni. Difazna nimetatakse võnkumiseks, kus kõrvalekalde kõver ühes suunas isoelektrilisest joonest ületab ja teostab võnkumist vastupidises etapis; Kolmefaasiline võnkumine teostab vastavalt kolm kõrvalekaldet isoelektrilise liini vastaskülgedel. Polüfhaas on võnkumine, mis sisaldab nelja või enama faasi.
Stimuleerimismeetodid elektromüograafias
Lisaks uurimisele elektrilise aktiivsuse lihaste üksi, refleksi ja suvalise lühenditega kaasaegne keeruline tehnika kliinilise elektromüograafia hõlmab uuring elektriliste reaktsioonide närvide ja lihaste elektrilise stimulatsiooni. Elektrienergia stimuleerimisest põhjustatud seadmed ja meetodid on samad, mis tavapärases elektromüograafias. Elektrostimulaatorid kasutatakse närvide ja lihaste stimuleerimiseks. Lihaste stimulatsiooni tehakse kodanikuelektroodid mootorpunktides, närvide stimulatsiooni vastavalt nende projektsiooni projektsiooni tsoonidele. Elektroodide stimuleerivad metallkettade kujul, mille läbimõõt on 6-8 mm, monteeritud metallklipi ja naatriumkloriidi isotoonilise lahuse märgamine. Neuromuskulaarsete haiguste diagnoosimise stimuleerimismeetodid lahendavad järgmisi peamisi ülesandeid: 1) lihaste otsese erutuse uurimine; 2) neuromuskulaarse ülekande uurimine; 3) uuring olukorra kohta Motiononons ja nende aksonid; 4) perifeersete närvide tundlike kiudude uurimine. Elektromüograafika abil on võimalik tuvastada, kas elektrienergia muutus on ühendatud mootori menüüsi või sünaptilise ja üle-vaimse struktuuri kahjustustega.
Elektromiograafilisi andmeid kasutatakse laialdaselt patoloogiliste või regeneratiivsete protsesside aktuaalse diagnoosi ja objektiivsete diagnooside selgitamiseks. Selle meetodi kõrge tundlikkus, mis võimaldab identifitseerida närvisüsteemi subkliinilisi kahjustusi, muudab selle eriti väärtuslik. Elektromüograafikat kasutatakse laialdaselt mitte ainult neuroloogilises praktikas, vaid ka teiste süsteemide lüüasaamise uurimisel, kui teiseks tõttu kahjustatud mootori funktsiooni (kardiovaskulaarne, vahetus, endokriinsed haigused).
Lihaste suvalise lõõgastumisega kogutakse ainult väga nõrk (kuni 10-15 μV) ja sagedased biopotentsiaal võnkumised. Lihastoonis refleksi muutusi iseloomustab väikese suurenemine sagedaste, kiirete ja lenduvate kõikuvate kõikumiste amplituudide suurenemisega biopotentsiaalides (kuni 50 μV). Lihaste meelevaldsete lühenditega registreeritakse interferentsi elektromogrammi (sagedaste kõrgepinge-biopotentsiaalidega kuni 2000 μV).
Sülitava seljaaju eesmise sarvede lüüasaamine põhjustab EMG-i muutuse sõltuvalt kahjuri raskusest, haiguse käigu olemusest ja selle etapist. Paraeside korral kokku lepitud rütmiliste võnkumiste kasv kestus kuni 15-20 ms täheldatakse. Kaitsekatuse või perifeerse närvi kahjustus põhjustab biopotentsiaalide amplituudi ja sageduse vähenemise, muutust EMG-kõvera kujul. Slugged paralüüs avaldub "bioelektrilise vaikuse".
Inimese käe ühe lihaste EMG on normaalne. . Elektromogrammi seljaaju esiklambrite lüüasaamist.

Küsimused üliõpilaste sõltumatu koolivälise töö jaoks:
Mootori seadme koosseis. Mootori basseini mõiste.
Mootori üksuste klassifikatsioon.
Rapid ja aeglase mootori osakute võrdlevad omadused.
Tervikliku lihase vähendamise tugevuse reguleerimine. Mootori osakute osaluse põhimõtted, mootori basseini fraktsioneerimine, ühine lõpptee.
Elektromüograafika meetod, meetodi põhimõte, EMG-meetodi meditsiiniline tähendus.
Praktilise töö sülearvutil valmistatakse ette EMG-meetodi lühikirjeldus (meetodi põhimõte, vajalikud seadmed, elektroodide liigid ja nende kasutamise omadused, meetodi meditsiiniline väärtus).